Human anatomy is undoubtedly a major core subject for study in medical schools. Despite the fact that normal human anatomy is a discipline that stood at the origins of the development of medicine, a large number of scientific works still appear that make adjustments to modern anatomical atlases.
It would seem that human anatomy cannot change so quickly with the course of evolution, but our understanding of it is constantly improving as new research methods appear - evidence of this is provided by ever new versions of the anatomy atlas.
Atlas of anatomy Sinelnikova R.D. in 4 volumes, this is perhaps the most authoritative and time-tested source of knowledge on this topic. It is constantly republished, delighting us with its visual illustrations and text accessible to everyone. Many students tried to download Sinelnikov’s atlas for study, but the links either did not work, or there was a virus in the folder... We solved this problem by making a website dedicated to this source.
The main goal of studying human anatomy is to create a fundamental knowledge base for students for further study of other medical disciplines. It is difficult to imagine mastering a curriculum in physiology, pathological physiology, pathological and topographic anatomy, operative surgery, and a whole range of clinical disciplines without a thorough study of normal human anatomy.
It is very important for a student to have a visual image of the material studied; for this it is necessary to study human anatomy in pictures. The main feature of this science. of course, is the structuring of its sections and subsections, as well as a clear systematization of the entire nomenclature.
Thus, we can distinguish the following directions that correspond to each system:
- osteology (section on the bones of the human skeleton). Studies the skeleton, both as a whole mechanism and as individual bones. There is also a study of age-related changes in bones.
- syndesmology (joints, ligaments). An extremely important section for future orthopedists and traumatologists.
- myology (muscular system). He studies not only the structure, but also the development and physiology.
- splanchnology (internal organs). Includes the anatomy of the endocrine, digestive, respiratory, excretory and genitourinary systems.
- angiology (vessels and their derivatives). Information is presented on the structure of blood and lymphatic vessels.
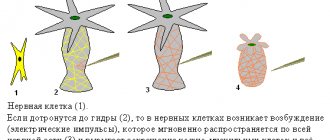
- neurology (central and peripheral nervous system). An extremely important section for successful diagnosis of diseases and perhaps the most difficult.
- aesthesiology (the science of the sense organs). Everything about vision and hearing. And also about taste, olfactory and tactile sensitivity. Closely related to neurology.
Text of the book “The Complete Course in 3 Days. Human anatomy"
Internal structure
On a cross-section of the bridge, three parts are distinguished: the ventral part (or base of the bridge) - the basilar part, the dorsal part, or the tegmentum of the bridge; and the trapezoidal body located between them.
The base of the bridge is formed by gray and white matter. The gray matter of the base of the bridge is represented by numerous small intrinsic nuclei of the bridge. The neurons of these nuclei are approached and synapse on them by nerve fibers originating from the neurons of the cerebral cortex - the corticopontine tract. The axons of the neurons of these nuclei are directed to the opposite side and, as part of the middle cerebellar peduncles, reach the cortex of the cerebellar hemispheres. This section of the pathway is called the “pontine-cerebellar tract.”
The white matter of the base of the bridge is represented by fibers having longitudinal and transverse directions. The longitudinally oriented corticospinal tract, which within the pons is represented by scattered bundles of fibers, passes through in transit. Only in the area of the lower edge of the bridge do they gather into a compact bundle and then form the pyramids of the medulla oblongata. The fibers of the corticonuclear tract have the same longitudinal direction at the base of the bridge, which partially ends on the neurons of the motor nuclei of the V, VI and VII pairs of cranial nerves, and then continues to the motor nuclei of the IX–XII pairs of cranial nerves located in the medulla oblongata. The dorsal part of the pons - its tire - also contains gray and white matter. The gray matter of the tegmentum is represented by the nuclei of the V, VI, VII and VIII pairs of cranial nerves, which are located mainly in the dorsolateral part of the tegmentum. In addition, there are nuclei that are part of the reticular formation, located on both sides of the median plane.
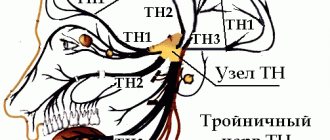
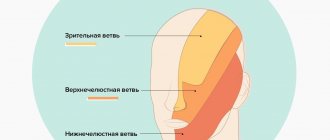
Trigeminal nerve, n. trigeminus, the V pair of cranial nerves, has nuclei not only within the pons. They are also located in the midbrain and spinal cord. The pontine tegmentum contains the pontine sensory nucleus, or superior sensory nucleus, and the motor nucleus of the trigeminal nerve. The pontine nucleus of the trigeminal nerve is the communication center of the tactile sensitivity pathway from the facial area. The sensory nucleus of the spinal tract of the trigeminal nerve is located in the dorsal horns of the four upper cervical segments (C1–C4) of the spinal cord. It is the communication center of the pain and temperature sensitivity pathway from the facial area. And finally, the V pair has a sensory nucleus in the midbrain - the nucleus of the midbrain tract, which is located in the central gray matter, lateral to the cerebral aqueduct. It plays the role of a communication center for the proprioceptive sensitivity of the facial muscles, palate muscles, neck muscles above the hyoid bone and the tensor tympani muscle.
Motor impulses to these muscles come from the motor nucleus, which is located in the pons tegmentum. All sensory nuclei of the trigeminal nerve are interneurons, the axons of which go to the integration centers of the brain as part of the nuclear-thalamic tract (the path of conscious impulses of general sensitivity from the head and neck area) and the nuclear-cerebellar tract (the path of unconscious proprioceptive impulses from the head and neck area). neck).
The first neurons of the afferent pathways are located in the trigeminal ganglion, ganglion trigeminal, which lies on the anterior surface of the pyramid of the temporal bone in the splitting of the dura mater at the trigeminal fusion, impressio trigemini.
Abducens nerve, n. abducens, the VI pair of cranial nerves, is a motor nerve in fiber composition. Its motor nucleus is located in the dorsal part of the pontine tegmentum. This nucleus is surrounded by arcuate fibers of the facial nerve, which form the facial tubercle in the upper part of the rhomboid fossa. The abducens nerve root emerges in the horizontal groove between the pons and the pyramids of the medulla oblongata.
Facial nerve, n. facialis, the VII pair of cranial nerves, is a mixed nerve in fiber composition, which has motor, sensory and parasympathetic nuclei. The axons of the cells of the motor nucleus are first directed dorsally, then arc around the motor nucleus of the abducens nerve and are directed ventrolaterally. The root of the facial nerve leaves the brain in the region of the cerebellopontine angle.
The parasympathetic nuclei of the facial nerve (superior salivary nucleus and lacrimal nucleus) are located in the lateral part of the pontine tegmentum. The axons of the cells of these nuclei leave the brain along with the fibers of the motor nucleus.
The sensory nucleus, in which the interneurons of the afferent pathways are located, is called the nucleus of the solitary tract. It is common to the X, IX and VII pairs of cranial nerves. Receptor neurons (pseudo-unipolar cells) associated with the facial nerve lie in the geniculate ganglion, which is located in the canal of the facial nerve.
The nuclei of the VIII pair of the vestibulocochlear nerve have complex architecture and connections. They are located on the border of the medulla oblongata and the pons in the lateral part of the rhomboid fossa, which is called the “vestibular-auditory field”.
Considering the complex structure of this nerve, it is advisable to separately consider its cochlear and vestibular parts.
It should be noted that the auditory nuclei lie in the most lateral part of the vestibular-auditory field and are represented by the ventral and dorsal cochlear nuclei. On the neurons of these nuclei, the central processes of the bipolar neurons of the spiral (cochlear) ganglion, located in the bony part of the cochlea, synaptically end. The totality of the central processes of bipolar cells makes up the cochlear (lower, or auditory) root of the vestibulocochlear nerve.
From the neurons of the cochlear nuclei, axons are directed to the nuclei of the trapezoid body. However, the course of the axons of the cells of the ventral and dorsal nuclei is significantly different. From the dorsal cochlear nucleus they extend onto the dorsal surface of the pons, forming the medullary (auditory) stripes of the fourth ventricle. The fibers then end on the neurons of the dorsal nucleus of the trapezoid body on the opposite side. The axons of the neurons of the ventral cochlear nucleus are directed to the neurons of the ventral nuclei of the trapezoid body of the opposite side. They pass in the ventral part of the bridge and form the basis of the trapezoid body. On a cross-section of the bridge, these fibers create transverse striations. The axons of the trapezoid body nuclei form a bundle of fibers called the lateral (auditory) lemniscus. These fibers are directed to the subcortical hearing centers (nucleus of the inferior colliculus of the midbrain, nuclei of the medial geniculate body and central nuclei of the thalamus optica). Along the lateral lemniscus, within the pons, there are nuclei of the lateral lemniscus, in which part of the fibers of the cochlear nuclei of the pons, passing in transit through the nuclei of the trapezoid body, are interrupted.
The vestibular nuclei lie somewhat laterally in relation to the cochlear nuclei. There are four nuclei on each side of the pons: the lateral vestibular nucleus, the medial vestibular nucleus, the superior vestibular nucleus, and the inferior vestibular nucleus. The central processes of bipolar cells located in the vestibular node end on the neurons of these nuclei. The latter lies at the bottom of the internal auditory canal. The totality of the central processes of the cells of the vestibular ganglion constitutes the vestibular (superior or vestibular) root of the vestibular-cochlear nerve. The axons of the cells of the vestibular nuclei form several bundles that go to the spinal cord, to the medial longitudinal fasciculus, to the cerebellum and to the central nuclei of the visual thalamus.
It should be noted that the gray matter of the pons is represented by four groups of nuclei:
1) the own nuclei of the bridge, located in its ventral part;
2) nuclei of the V–VIII pairs of cranial nerves, which are located in the tegmentum of the bridge;
3) the nuclei of the reticular formation, also located in the operculum of the bridge;
4) nuclei of the trapezoid body.
The white matter of the bridge tire consists of longitudinal fibers. Most of them are afferent and come here from the spinal cord and medulla oblongata. Near the lateral edge of the tegmentum there is a bundle of fibers of the anterior spinocerebellar tract. More medially is the spinothalamic tract, which unites here the fibers of the lateral and anterior spinothalamic tracts. Even more medial are bundles of nerve fibers starting in the nuclei of the medulla oblongata: the nuclear-thalamic tract and the bulbar-thalamic tract.
The most significant bundle of efferent fibers of the pontine tegmentum is the red nucleus-spinal cord, which is located in the lateral section. Near it is the vestibular spinal tract, the fibers of which originate mainly from the lateral vestibular nucleus, the VIII pair of cranial nerves.
A relatively small bundle begins from the nuclei of the reticular formation of the bridge, which joins the reticular-spinal cord tract, starting from the cells of the reticular formation of the diencephalon and midbrain.
Dorsally, near the midline, there are medial and posterior longitudinal fasciculi. Anterior (ventral) to the medial longitudinal fasciculus runs the tegmental spinal cord, which originates from the neurons of the superior colliculus of the midbrain.
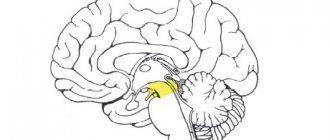
Cerebellum
The cerebellum, cerebellum, develops from the dorsal wall of the hindbrain and is the largest part of the brain after the cerebral hemispheres.
Together with the medulla oblongata and the pons, the cerebellum is located in the posterior cranial fossa.
The cerebellum has a rhomboid shape with a predominance of transverse size. Its width is on average 10 cm, length along the midline is 3–4 cm, thickness is 4–5 cm; average weight - about 135 g.
The cerebellum is divided into a middle part - the vermis, and two voluminous lateral parts - the hemispheres. Based on the development of the cerebellum in phylogeny, a small formation adjacent to the hemisphere on the ventral side, a flocculus, should be distinguished. In the vermis and cerebellar hemispheres, two surfaces are distinguished: superior and inferior.
The superior surface of the cerebellum faces upward and backward. It is convex and has a longitudinal elevation in the middle, called the superior vermis. The worm passes into the hemispheres from the lateral sides. The inferior surface of the cerebellum is directed downward and forward. It is adjacent to the occipital bone. On the lower surface of the cerebellum there is a longitudinal depression called the valley. This depression contains the inferior vermis, separated from each hemisphere by a groove.
The surface of the cerebellum is striated with a large number of grooves parallel to each other, which have a transverse direction and varying depths. Shallow grooves divide the surface of the cerebellum into plates, deeper grooves divide groups of plates into plates, which are called “lamellae of the cerebellum”. Finally, the deepest grooves divide the surface of the cerebellum into lobules.
Among the grooves separating the cerebellar lobules, the deepest (up to 2 cm) is the horizontal groove. It runs along the entire circumference of the cerebellum and separates the superior and inferior surfaces of the hemispheres.
The furrows of the cerebellum, without interruption, pass from the vermis to the hemispheres. The shred also has plates, but no segments are distinguished in it.
There are 8 lobules in the worm, 4 lobules in the upper and lower. The most anterior lobule of the superior vermis is the uvula. This lobule is formed by several plates. The next one is the central lobe, which corresponds to the upper, most protruding part of the worm. Posterior to the central lobule there is a hill, in which a front part (apex) and a back part (slope) are distinguished. The most posterior lobe of the upper vermis is the leaf of the vermis, which limits the horizontal groove at the top.
In the lower worm, below the leaf, there is a worm tubercle. Anterior to it is the pyramid of the vermis, protruding at the bottom of the cerebellar valley. Further anteriorly lies the narrowest part of the lower worm - the uvula of the worm, which is, as it were, compressed by the adjacent parts of the hemispheres. Finally, the most anterior lobule of the inferior vermis is the nodule.
In each hemisphere, the lobules of the worm correspond to the lobes of the hemispheres. On the upper surface of the hemisphere, the quadrangular lobule and the superior semilunar lobule are defined.
On the lower surface of the hemisphere, from back to front, are the inferior semilunar lobule, the thin lobule, the digastric lobule and the cerebellar amygdala.
The flocculus is a small group of cerebellar plates adjacent to its middle peduncle.
This division of the cerebellum into lobules was given on the basis of the presence of connections between individual parts of the hemispheres and certain areas of the vermis. Modern studies of the cerebellar pathways suggest that it is more rational to isolate parts whose function was formed in the process of phylogeny and ontogenesis. Thus, in the cerebellum there is a phylogenetically ancient part (paleocerebellum), which includes the flocculus and nodule, the old part (archicerebellum), which includes the vermis with the exception of the nodule, and the new cerebellum (neocerebellum), which includes the cerebellar hemispheres developing from the middle part of the vermis.
Internal structure
On the sections you can see the gray matter located on the surface, forming the cerebellar cortex, under the cerebellar cortex - the white matter, which makes up the brain body, from which processes extend to the surface, penetrating into the lobules and plates of the cerebellum. On the median section of the cerebellum, the white matter has a leaf-like shape, which is associated with the figurative name “tree of life.”
In the thickness of the white matter of the cerebellum there are accumulations of gray matter that make up the cerebellar nuclei. In the cerebellar vermis, on either side of the midline, there is a tent nucleus. Lateral to this nucleus there is a second small nucleus, called the spherical nucleus. The corky nucleus lies even more laterally. The white matter of the hemispheres contains the largest nucleus, the dentate nucleus.
The tent nucleus belongs to the ancient cerebellum, the globular and cortical nuclei are phylogenetically later formations (belong to the old cerebellum), and the dentate nucleus belongs to the new cerebellum.
The white matter of the cerebellum contains afferent and efferent fibers that connect the cerebellum to the brainstem and form the cerebellar peduncles. There are three pairs of cerebellar peduncles: superior, middle and inferior. The superior cerebellar peduncles connect it to the midbrain, the middle cerebellar peduncles to the pons, and the inferior cerebellar peduncles to the medulla oblongata. The superior and inferior cerebellar peduncles are visible from the dorsal surface of the brainstem, and the middle peduncles are visible from its ventral surface.
The cerebellum is fundamentally different in structure from other previously discussed parts of the brain (medulla oblongata and pons). This difference lies in the fact that the outside of the cerebellum is covered with a layer of gray matter - the cerebellar cortex, which is a collection of neurons, the location of which is strictly subject to a certain pattern. There are three layers in it: outer - molecular, middle - ganglionic, inner - granular. This layer-by-layer arrangement of neurons is a characteristic morphological feature of the integration centers of the brain, one of which is the cerebellum. This explains the numerous complex connections of the cerebellum with other parts of the central nervous system.
Connections between the cerebellum and the spinal cord and brain
The most ancient connections of the cerebellum were established with the organs of balance. Nerve fibers extend from the vestibular nuclei, representing part of the vestibular-cerebellar tract. They penetrate the cerebellum as part of its lower peduncles and end on the neurons of the flocculus and nodule cortex. The descending (efferent) pathway begins from the neurons of these areas of the cerebellar cortex. Nerve fibers from the flocculus and nodule cortex reach the neurons of the tent nucleus, which is the oldest of the cerebellar nuclei. The axons of the neurons of the tent nucleus reach the nuclei of the reticular formation of the medulla oblongata through the inferior cerebellar peduncles. From them, along the reticular-spinal cord tract, efferent impulses arrive to the muscles of the trunk.
Later, in connection with the emergence of animals on land and the development of limbs, afferent pathways from the receptors of muscles, tendons and joints of the limbs appeared in the form of the posterior and anterior spinocerebellar tracts. Fibers of the posterior spinocerebellar tract (Flexig's bundle) pass into the cerebellum as part of the inferior cerebellar peduncles and end on neurons of the cortex of the posterior part of the vermis. The fibers of the anterior spinocerebellar tract (Gowers' bundle) enter the cerebellum as part of its superior peduncles and end on the neurons of the cortex of the anterior part of the vermis. The efferent pathway from the neurons of the vermis cortex goes to the neurons of the globular and cortical nuclei of the cerebellum. The axons of the neurons of these nuclei exit the cerebellum as part of its superior peduncles and reach the neurons of the reticular formation. Subsequently, efferent impulses along the reticular-spinal cord tract are sent to the muscles of the limbs.
Due to the complication of the functions of the muscles of the trunk and limbs, the bulbar-cerebellar and nuclear-cerebellar pathways have also formed, which end in the cortex of the middle part of the cerebellar vermis. The efferent pathway from the neurons of the middle part of the cortex of the vermis is similar to the previous ones.
In mammals and especially in humans, a system of so-called precerebellar nuclei has developed: the olivary nuclei and the proper pontine nuclei. The olive nuclei receive impulses through collaterals extending from the fibers of the extrapyramidal tract. Axons from olivary neurons pass through the inferior cerebellar peduncles to form the olivocerebellar tract. The fibers of this pathway pass in the medulla oblongata to the opposite side and end on the neurons of the cerebellar cortex.
The pontine nuclei are communication centers along the corticopontine-cerebellar pathway. The neurons of these nuclei terminate the fibers of the cortical-pontine tracts, as well as the collaterals of the pyramidal fibers. The axons of the neurons of the own nuclei of the pons move to the opposite side and, as part of the middle cerebellar peduncles, are directed to the neurons of the cortex of the cerebellar hemispheres. From the cortex of the cerebellar hemispheres, the cerebellar-dentate-rednuclear-spinal cord tract begins, which carries out the so-called corrective activity when performing complex voluntary movements, mainly with the upper limb.
The complex neural connections of the cerebellum with the cerebral cortex explain the diversity of its functions. The cerebellum is involved in coordinating the work of various muscle groups (flexors and extensors), helps overcome the inertia of rest and inertia of movement (rapid alternation of pronation and supination of hands extended forward), and ensures the accuracy of fine movements (finger-to-nose test). Damage to the structures of the cerebellum is accompanied by a violation of its characteristic functions, which are manifested by symptoms such as cerebellar ataxia (drunk gait), intentional tremor when performing fine movements and impaired coordination of movements.
It should be noted that the efferent pathways of the cerebellum intersect twice (before the red nucleus and after the red nucleus). As a result of this, unilateral damage to the cerebellum is manifested by loss of function of the muscles of the same side. Even unilateral cerebellar lesions cause imbalance in the body.
IV ventricle
The fourth ventricle is the cavity of the rhombencephalon. It is a continuation of the central canal of the spinal cord. The bottom of the IV ventricle is formed by a rhomboid fossa, which is truly diamond-shaped, bounded by the superior and inferior cerebellar peduncles. It has two halves: lower (caudal) and upper (rostral), delimited by medullary stripes. The lower half of the rhomboid fossa is the dorsal surface of the medulla oblongata, the upper half is the dorsal surface of the pons.
A median groove runs along the midline of the rhomboid fossa, on both sides of which there is a longitudinally shaped medial elevation. Laterally it is limited by the border groove. This groove is of great importance, as it serves as an approximate boundary between the projection of the motor and sensory nuclei of the cranial nerves. The motor nuclei project more medially, the sensory nuclei project laterally. The medial eminence at the lower corner of the rhomboid fossa is called the triangle of the hypoglossal nerve. Above the stria medullaris, the medial eminence forms a noticeable thickening called the facial tubercle.
The roof of the IV ventricle has two parts that differ in development and structure. In the region of the medulla oblongata, the dorsal wall of the neural tube atrophies and remains in the form of ependymal epithelium. A duplication of the choroid of the fourth ventricle is adjacent to the outer surface of the epithelial plate.
The lower (posterior) cerebral velum is paired. It is stretched between the worm's nodule, the shred's leg and the shred.
In the area of the pons, the cerebellum develops from the dorsal wall of the neural tube. The anterior part of the roof of the IV ventricle is formed by a plate of white matter - the superior (anterior) medullary velum, which is stretched between the superior cerebellar peduncles.
Thus, the posterior part of the roof of the IV ventricle is represented by the posterior medullary velum and the pia mater. The latter is fused with the free edge of the posterior medullary velum, with the lower cerebellar peduncles and the posterior cords of the spinal cord.
The cavity of the fourth ventricle communicates below with the central canal of the spinal cord, and above with the aqueduct of the midbrain. In addition, in the region of the lateral angles of the rhomboid fossa in the vascular base of the IV ventricle there is a connection with the subarachnoid interthecal space through a paired opening called the lateral aperture. There is another unpaired foramen in the region of the lower corner of the rhomboid fossa - the median aperture (foramen of Mozhandi). Through these openings, cerebrospinal fluid flows from the fourth ventricle into the subarachnoid interthecal space of the brain.
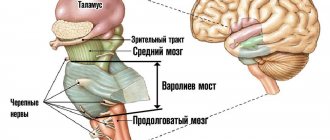
Midbrain
The midbrain, mesencephalon, develops from the midbrain. Compared to other sections, the midbrain is small in size. Its ventral surface is represented by the cerebral peduncles and the posterior perforated substance located between them. The dorsal surface is formed by the roof of the midbrain; the cavity is the aqueduct of the midbrain (Aqueduct of Sylvius).
On the ventral side, the cerebral peduncles look like two thick, flattened ridges that appear from under the upper edge of the pons. From here they are directed upward and to the sides at an angle of 70–80° and are immersed in the substance of the diencephalon. The anterior border of the cerebral peduncles is the optic tract, which belongs to the diencephalon. The cerebral peduncles are white in color and have a fibrous structure due to the longitudinal arrangement of nerve fibers. The cerebral peduncles have a base and a tegmentum.
Along the medial edge of the base of the cerebral peduncles there is a groove of the oculomotor nerve, from which the oculomotor nerve, the third pair of cranial nerves, emerges as one root.
Along the lateral edge of the cerebral peduncle there is a lateral fissure of the midbrain, which is a continuation of the fissure of the rhombencephalon, separating the superior and middle cerebellar peduncles.
On the ventral side between the two cerebral peduncles there is a depression called the interpeduncular fossa. It is narrower, at the upper edge of the bridge it expands anteriorly and ends near the two papillary bodies belonging to the diencephalon. The surface of the interpeduncular fossa has a grayish color and is dotted with holes through which numerous blood vessels pass. This area of the brain is called the posterior perforated substance.
On the dorsal surface of the midbrain, represented by the roof plate, there are four rounded elevations - two superior colliculi and two inferior colliculi. The mounds are separated by grooves intersecting at right angles. The lower hillocks are smaller than the upper ones.
The handles of the hillocks extend from each hillock on the lateral side. They move forward and upward to the diencephalon. The handles of the superior colliculi, narrower and longer, end in the lateral geniculate bodies. The handles of the inferior colliculi, thicker and shorter, end in the medial geniculate bodies.
Posterior to the inferior colliculi along the midline is the frenulum of the superior medullary velum, which has a triangular shape. On each side of the frenulum of the superior medullary velum, one root of the fourth pair of cranial nerves emerges. Trochlear nerve, n. trochlears, IV pair of cranial nerves - the thinnest of all cranial nerves and the only nerve that exits the substance of the brain on its dorsal surface. The nerve then bends around the cerebral peduncles and goes to their ventral surface.
On the lateral surface of the midbrain, in the interval between the lateral sulcus of the midbrain and the handles of the inferior colliculi, a triangular-shaped area is distinguished - the loop triangle. The third side of the triangle is the lateral edge of the superior cerebellar peduncle. In the projection of the triangle, in the thickness of the cerebral peduncles, nerve fibers pass that make up the lateral, medial, trigeminal and spinal lemniscus. Thus, in this place, in a small area near the surface of the brain, almost all the pathways of general sensitivity (conducting impulses to the diencephalon) and the auditory pathway are concentrated. In neurosurgical practice, cordotomy is performed within the loop triangle - an operation for intolerable pain due to lesions of the visual thalamus.
The cavity of the midbrain is the midbrain aqueduct. It is a remnant of the midbrain cavity. The aqueduct is oriented along the axis of the brain, connecting the IV and III ventricles. Its length is about 15 mm, average diameter is 1–2 mm. There is a slight expansion in the middle part of the cerebral aqueduct. The beginning of the aqueduct is located in the upper corner of the IV ventricle. It is covered by the superior cerebral velum. The hole through which the aqueduct opens into the third ventricle is located under the posterior commissure of the cerebrum.
Internal structure
A cross-section of the midbrain clearly identifies its main parts: above the aqueduct there is a plate of the roof, below there are the cerebral peduncles. A section through the cerebral peduncles reveals a pigmented layer of gray matter called the substantia nigra (Semmering's substance). The substantia nigra delimits the base of the cerebral peduncles and the tegmentum of the midbrain.
The base of the cerebral peduncles is formed mainly by longitudinally oriented descending fibers that go from the neurons of the cerebral cortex to the nuclei of the brain stem and spinal cord. In this regard, the base of the cerebral peduncles is a phylogenetically new formation.
On a cross section, the substantia nigra has the shape of a flattened crescent with a convexity facing ventrally. The dorsal part of the substantia nigra contains highly pigmented nerve cells containing large amounts of iron. The ventral part of the substantia nigra contains large scattered nerve cells and myelin fibers passing between them.
The tegmentum of the midbrain contains gray and white matter. The gray matter is represented by the paired red nucleus and the central gray matter located around the cerebral aqueduct.
The red nuclei are cylindrical in shape, located throughout the midbrain in the center of the tegmentum of each cerebral peduncle and continue into the diencephalon.
The caudal part of the red nucleus contains large nerve cells, while the rostral part contains mainly small nerve cells. The cells of the red nucleus, like the cells of the substantia nigra, contain iron, but in much smaller quantities. The neurons of the red nucleus terminate in the fibers of the dentate-red nuclear tract and the axons of the cells of the basal nuclei of the telencephalon, forming the striatal-red nuclear tract. The axons of neurons of large cells of the red nucleus unite into the red nucleus-spinal cord tract. The axons of small neurons of the red nucleus end on the neurons of the reticular formation and olives of the medulla oblongata, forming the red nuclear-reticular and red nuclear-olive tracts.
Ventral to the red nucleus is the unpaired interpeduncular nucleus. The neurons of this nucleus terminate in fibers originating from the leash nuclei. The latter are located in the epithalamus of the diencephalon. The bundle of fibers starting from the leash nuclei is called the “lead-interpeduncular path.” It can be assumed that this bundle is one of the links in the efferent autonomic pathway.
Surrounding the cerebral aqueduct is the central gray matter. In the ventrolateral part of this substance, at the level of the inferior colliculi, the motor nuclei of the fourth pair of cranial nerves - the trochlear nerve - are located. The axons of the neurons of these nuclei are directed dorsally, pass to the opposite side and exit the brain substance in the region of the frenulum of the superior medullary velum. Cranial to the motor nuclei of the IV pair of cranial nerves (at the level of the superior colliculi) are the nuclei of the III pair of cranial nerves - the oculomotor nerve.
The oculomotor nerve has three nuclei. The motor nucleus is the largest and has an elongated shape. It has five segments, each of which provides innervation to certain muscles of the eyeball, and the muscles that lift the upper eyelid.
The segments, if we consider the nucleus from top to bottom, innervate the following muscles:
1) the muscle that lifts the upper eyelid;
2) superior rectus muscle;
3) inferior oblique muscle;
4) inferior rectus muscle;
5) medial rectus muscle.
In addition to the paired motor nucleus, the oculomotor nerve has another unpaired nucleus - the central unpaired nucleus. This nucleus is interconnected with the caudal segments of the main motor nuclei of both sides, which are responsible for the innervation of the medial rectus muscles. This ensures the combined work of the indicated muscles of the right and left eyeballs, which rotate the eyeball and bring the pupils closer to the median plane. Due to its function, the central unpaired nucleus is also called convergent.
Dorsal from the motor nuclei, near the midline, there is a paired autonomic nucleus of the oculomotor nerve - the so-called accessory nucleus (Yakubovich's nucleus). The neurons of this nucleus are responsible for the innervation of the constrictor pupillary muscle and the ciliary muscle.
Some fibers from the motor somatic nuclei of the oculomotor nerve participate in the formation of the medial longitudinal fasciculus. Most of the fibers from all nuclei make up the root of the oculomotor nerve, which emerges from the substance of the brain in the groove of the same name.
In the lateral part of the central gray matter is the nucleus of the midbrain tract of the trigeminal nerve (midcerebral nucleus).
Between the central gray matter and the red nuclei is the reticular formation, which contains numerous small nuclei and two large nuclei. One of them is called the intermediate nucleus (nucleus of Cajal), the second is the nucleus of the posterior commissure of the brain (nucleus of Darkshevich). The axons of the cells of the Cajal nucleus and the Darkshevich nucleus are sent to the spinal cord, forming the medial longitudinal fasciculus.
Possible pathologies and their diagnosis
The significance of the bridge can be assessed based on the influence of pathologies (syndromes) that damage individual functions of the body.
The most common of them include:
- Bonnier syndrome is accompanied by damage to the nuclei of the auditory and vestibular nerves. In this case, the patient becomes dizzy, hearing loss occurs, and trigeminal neuralgia may occur. Common symptoms include weakness, depression, and sleep disturbances.
- Locked-in syndrome (ventral pontine syndrome) is a condition in which consciousness and full sensitivity are retained, but the ability to speak is completely lost. The function of the extraocular muscles is preserved. Communication with others is possible using non-verbal gestures. The condition is preceded by signs confirming insufficiency of the blood supply artery: double vision, dizziness, unsteadiness of gait.
- Raymond-Sestan syndrome (another name is oral tegmentum syndrome) is a combination of paralysis of the muscles responsible for the movement of the eyeball on the side opposite to the lesion. Etiological factors: atherosclerotic changes in cerebral vessels, tumors, ischemic strokes.
- Millard-Hubler syndrome is manifested by paralysis of the facial muscles on the affected side, along with partial paralysis on the opposite side. This disease manifests itself in pathologies at the base of the bridge. Constriction of blood vessels or micro-stroke predisposes to pathology, for example, if there is a cavernous angioma in a given structure with subsequent damage to the structures of the vascular system. Less commonly, the cause may be neurosyphilis or diffuse glioma.
- Foville syndrome is a combined lesion of individual elements of the facial and abducens nerves. The pathology is expressed in complete paralysis of the facial muscles in combination with strabismus. Often the cause of its development is ischemic stroke, less often tumor-like formations, inflammation.
- Gasperini syndrome is caused by the occurrence of pathology in the area of the bridge tire. It affects the nuclei of several nerves at once (facial, trigeminal, vestibulocochlear abducens). From the location of the pathological focus on the opposite side, a person feels a sensitivity disorder. The clinical picture includes strabismus, dizziness, and ataxia. This condition occurs due to ischemia, tumors, and inflammation.
- Grenet's syndrome is a sensory disorder with simultaneous damage to the muscles responsible for chewing on the affected side. On the opposite side, hemihypesthesia is noted. Often, pathology can occur due to ischemic changes in the branches of the posterior cerebral artery.
- Brissot-Sicart syndrome is a set of signs of damage to the nucleus of the facial nerve with partial paralysis of the limbs. Clinically manifested by a spasm of the facial muscles, which is accompanied by peripheral paralysis of the facial nerve and hemiparesis. Its occurrence is associated with ischemia and previous infectious diseases.
Important Sermion: cheap analogues and substitutes, prices for Russian and foreign drugs in tablets
Modern methods of magnetic resonance imaging help to clarify the location, duration of the lesion, volume and other parameters of the pathological process.
Like any organ in the human body, the VM can also stop functioning and this can be caused by the following diseases:
- stroke of the cerebral arteries;
- multiple sclerosis;
- head injuries. Can be obtained at any age, including during childbirth;
- tumors (malignant or benign) of parts of the brain.
In addition to the main reasons that can provoke brain pathologies, you need to know the symptoms of such a lesion:
- the process of swallowing and chewing is impaired;
- loss of skin sensitivity;
- nausea and vomiting;
- nystagmus is eye movements in one specific direction, as a result of such movements one can often feel dizzy, even to the point of loss of consciousness;
- may see double when turning the head sharply;
- disturbances in the functioning of the motor system, paralysis of certain parts of the body, muscles, or tremors in the hands;
- in case of disturbances in the functioning of the facial nerves, the patient may experience complete or partial anemia, lack of strength in the facial nerve;
- speech disorders;
- asthenia – decreased strength of muscle contraction, rapid muscle fatigue;
- dysmetria - incompatibility between the task of the movement being performed and muscle contraction, for example, when walking, a person may raise his legs much higher than necessary or, on the contrary, may stumble over small bumps;
- snoring, in cases where it has never been observed before.