Vision is one of the basic senses of a person, which affects almost all aspects of life: orientation in space, obtaining information about surrounding objects, learning and entertainment, recognizing the emotions of other people, and much more.
The role of the control center of this sense is performed by the visual cortex of the brain. Many vision diseases occur due to disturbances in the functioning of this area. Therefore, to find methods for treating these diseases, much attention is paid to studying the structure and functions of the visual cortex.
General information
The visual cortex is a region of the cerebral cortex that receives, analyzes and processes visual information about the world around us.
It is closely connected with associative areas that are responsible for higher nervous functions. This area is located in the occipital and medial region of both hemispheres and occupies the 17th, 18th and 19th Brodmann cytoarchitectonic areas.
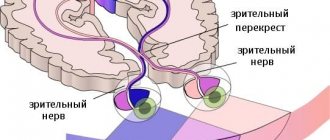
Visual data from the left half of the optic stream is received by the parts of the cortex of the right hemisphere, and the right by the left.
Sense organs (analyzers). Structure and functions of the organs of vision and hearing
Sense organs (analyzers). Structure and functions of the organs of vision and hearing
Analyzer
- a functional system consisting of:
- receptor,
- sensitive pathway
- the corresponding zone of the cortex where this type of sensitivity is projected.
Analysis and synthesis of the received information is carried out in a strictly defined area - the cerebral cortex zone
.
Based on the characteristics of the cellular composition and structure, the cerebral cortex is divided into a number of areas called cortical fields
.
The functions of individual areas of the cortex are not the same. Each receptor apparatus on the periphery corresponds to an area in the cortex - the cortical nucleus of the analyzer.
The most important zones of the cortex are as follows:
• Motor zone
located in the anterior central and posterior central regions of the cortex (anterior central gyrus in front of the central sulcus of the frontal lobe).
• Sensitive area
(the area of musculocutaneous sensitivity is located behind the central sulcus, in the posterior central gyrus of the parietal lobe). The largest area is occupied by the cortical representation of the receptors of the hand and thumb, vocal apparatus and face, the smallest area is occupied by the representation of the trunk, thigh and lower leg.
• Visual area
concentrated in the occipital lobe of the cortex. It receives impulses from the retina and distinguishes between visual stimuli.
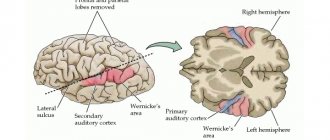
• Auditory zone
located in the superior temporal gyrus of the temporal lobe.
• Olfactory and gustatory zones
- in the anterior section (on the inner surface) of the temporal lobe of each hemisphere.
The activity of the cerebral cortex of humans and higher animals was determined by I.P. Pavlov as higher nervous activity
, which is a conditioned reflex function of the cerebral cortex.
Analyzers
are a set of nerve formations that provide awareness and assessment of stimuli acting on the body. The analyzer consists of receptors that perceive irritation, a conductive part and a central part - a certain area of \u200b\u200bthe cerebral cortex where sensations are formed.
Visual analyzer
provides visual information from the environment and consists of three parts:
peripheral - the eyes, conductive - the optic nerve, central - subcortical and visual zone of the cerebral cortex.
Eye
consists of the eyeball and auxiliary apparatus, which includes the eyelids, eyelashes, lacrimal glands and muscles of the eyeball.
The eyeball
is located in the orbit and has a spherical shape and 3
membranes
:
fibrous ,
the posterior part of which is formed by an opaque
tunica
albuginea (
sclera
),
choroid, retina
. Part of the choroid, equipped with pigments, is called the iris.
In the center of the iris there is
a pupil
, which can change the diameter of its opening due to contraction of the eye muscles.
Posterior retina
perceives light stimuli.
Its front part
is blind and does not contain photosensitive elements.
The photosensitive elements
of the retina are:
sticks
(provide vision in twilight and darkness),
cones
(color vision receptors that work in high light).
Cones are located closer to the center of the retina (macula macula), and rods are concentrated at its periphery. The exit point of the optic nerve is called the blind spot
.
The cavity of the eyeball is filled with vitreous humor
.
Lens
has the shape of a biconvex lens.
It is able to change its curvature when the ciliary muscle contracts. When viewing close objects, the lens contracts, and when viewing distant objects, it expands. This ability of the lens is called accommodation
.
Between the cornea and the iris is the anterior chamber of the eye
, between the iris and the lens is
the posterior chamber
.
Both chambers are filled with a clear liquid. Rays of light, reflected from objects, pass through the cornea, moist chambers, lens, vitreous body and, thanks to refraction in the lens, fall on the macula
of the retina - the place of best vision.
In this case, a real, inverse, reduced image of the object
.
From the retina, along the optic nerve, impulses enter the central part of the analyzer - the visual zone of the cerebral cortex
located in the occipital lobe. In the cortex, information received from retinal receptors is processed and a person perceives the natural reflection of an object.
Normal visual perception
due to:
– sufficient luminous flux;
– focusing the image on the retina (focusing in front of the retina means myopia, and behind the retina means farsightedness);
– implementation of the accommodative reflex.
The most important indicator of vision
is its sharpness, i.e.
the ultimate ability of the eye to distinguish small objects. Accommodation is
the adaptation of the eye to seeing objects at different distances.
During accommodation, muscles contract, which change the curvature of the lens. With constant excessive curvature of the lens, light rays are refracted in front of the retina and as a result, myopia .
If the curvature of the lens is insufficient, then light rays are focused behind the retina and
farsightedness occurs.
Myopia develops when the longitudinal axis of the eye is enlarged. Parallel rays coming from distant objects are collected (focused) in front of the retina, which is hit by diverging rays, resulting in a blurry image. For myopia, glasses with diverging biconcave glasses are prescribed, which reduce the refraction of rays so much that the image of objects appears on the retina. Farsightedness occurs when the axis of the eyeball is shortened. The image is focused behind the retina. To correct vision, biconvex glasses are required. Senile farsightedness usually develops after 40 years, when the lens loses elasticity, hardens and loses the ability to change curvature, which makes it difficult to see clearly at close range. The eye loses the ability to clearly see objects at different distances.
Organ of hearing and balance.
Hearing analyzer
ensures the perception of sound information and its processing in the central parts of the cerebral cortex.
The peripheral part
of the analyzer is formed by the inner ear and the auditory nerve.
The central part
is formed by the subcortical centers of the midbrain and diencephalon and the temporal zone of the cortex.
The ear
is a paired organ consisting of:
The outer ear
- includes the pinna, external auditory canal and eardrum.
The middle ear
consists of the tympanic cavity, the chain of auditory ossicles and the auditory (Eustachian) tube.
The auditory tube connects the tympanic cavity with the nasopharynx cavity. This ensures equalization of pressure on both sides of the eardrum. The auditory ossicles - the hammer, incus and stapes - connect the eardrum with the membrane of the oval window leading to the cochlea. The middle ear transmits sound waves from a low-density environment (air) to a high-density environment (endolymph), which contains the receptor cells of the inner ear. Inner ear
- located in the thickness of the temporal bone and consists of a bony labyrinth and a membranous labyrinth located in it.
The space between them is filled with perilymph, and the cavity of the membranous labyrinth is filled with endolymph. The bony labyrinth is divided into three sections - the vestibule, cochlea and semicircular canals
.
The organ of hearing includes the cochlea
- a spiral canal of 2.5 turns.
The cochlear cavity is divided by a membranous main membrane consisting of fibers of different lengths. On the main membrane are receptor hair cells
. Vibrations of the eardrum are transmitted to the auditory ossicles. They amplify these vibrations almost 50 times and are transmitted through the oval window into the fluid of the cochlea, where they are perceived by the fibers of the main membrane. The receptor cells of the cochlea perceive irritation coming from the fibers and transmit it along the auditory nerve to the temporal zone of the cerebral cortex. The human ear perceives sounds with a frequency from 16 to 20,000 Hz.
Organ of balance
or
the vestibular apparatus
is formed by two
sacs
and
three semicircular canals
.
Receptor hair cells
are located on the bottom and inside of the sacs. Adjacent to them is a membrane with crystals - otoliths containing calcium ions. The semicircular canals are located in three mutually perpendicular planes. At the base of the canals are hair cells. Receptors of the otolithic apparatus respond to acceleration or deceleration of rectilinear movement. The semicircular canal receptors are stimulated by changes in rotational movements. Impulses from the vestibular apparatus travel through the vestibular nerve to the central nervous system. Impulses from receptors in muscles, tendons, and soles also come here. Functionally, the vestibular apparatus is connected with the cerebellum, which is responsible for coordination of movements and orientation of a person in space.
Taste analyzer
consists of receptors located in the taste buds of the tongue, a nerve that conducts impulses to the central section of the analyzer, which is located on the inner surfaces of the temporal and frontal lobes.
Olfactory analyzer
represented by olfactory receptors located in the nasal mucosa. Along the olfactory nerve, the signal from the receptors enters the olfactory zone of the cerebral cortex, located next to the taste zone.
Skin analyzer
consists of receptors that perceive pressure, pain, temperature, touch, pathways and a zone of skin sensitivity located in the posterior central gyrus.
Anatomy of vision structure
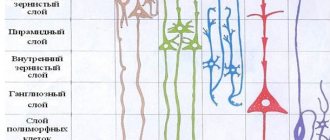
Like all other areas of the brain responsible for the senses, this area is divided into sensory and associative areas. The sensory area is called the primary visual cortex (striate or area V1), and the associative area is called the secondary visual cortex (prestriate).
Zone V1 is located in the area of the calcarine groove, and is stretched along the medial side. Its position corresponds to the 17th Brodmann field. Like almost the entire neocortex, it consists of 6 horizontal layers, but layer IV has significant differences from other areas. Layer IV in this case is divided into four sublayers. Division occurs by Jenari stripes, which consist of myelin fibers. In addition, there is a theory about the division of the remaining layers into a larger number of sublayers, but to date, this hypothesis has not been confirmed.
The prestriate cortex consists of small association areas that lie away from the central plane and surround area V1. At the moment, there are about 30 different associative areas, which are designated by the Latin letter V and a number after it, indicating the distance from the primary one. The first section, which lies close to zone V1, is designated V2. All visual association areas correspond to Brodmann's areas 18 and 19.
The structure of the areas of the prestriate cortex is almost identical to the entire neocortex. They also have 6 horizontal layers and rows of vertical columns. The only difference is that in these areas, layer VI has a more rich and complex architecture.
A person without a visual cortex is able to associate sound and visual stimulus
Rice. 1.
Pathways for transmitting visual information in the brain. Th (thalamus) - thalamus; V1 (visual 1) - primary visual cortex; SC (superior colliculus) - one of the two superior colliculi; Pulv (pulvinar) - pillow; LGN (lateral geniculate nucleus) - lateral geniculate body. The figure also shows the ventral and dorsal pathways for information transmission in the cortex; they are described in more detail in the news The brain sorts visual images differently than previously thought (“Elements”, 08/23/2013). Image from N. Diederich et al., 2014. Are patients with Parkinson's disease blind to blindsight?
We need the cerebral cortex (neocortex) in order to establish connections between information received through the senses. In the neocortex, images of objects are formed (for example, a banana is not only yellow, but also smooth), and it was thanks to it that Pavlov’s dog was able to understand that food does not appear on its own, but only after turning on the light bulb. However, new studies of neurological patients show that associative learning is possible without the participation of the neocortex. Thus, a person who has lost the visual cortex can establish an association between a circle of a certain color and size and a sound of a certain pitch and volume - and this despite the fact that he does not consciously perceive the colors, sizes and shapes of objects.
The phenomenon of pseudo-blindness, or blindsight (see Blindsight), has been known for several decades. And in recent years, its popularization has been greatly promoted by the science fiction novel of the same name by Peter Watts, a specialist in marine biology (see “Blindsight”, “False Blindness”) In blindsighted people, everything is fine with the eyes, but the work of the primary visual cortex of the cerebral hemispheres is disrupted. Therefore, such people do not distinguish the colors and shapes of objects, and also do not understand whether these objects are moving or not. Nevertheless, blindsighted people are able to move independently, avoiding obstacles, and can describe the object standing in front of them. And although the subjects claim that they do not see any object, but only guess its characteristics, the percentage of correct answers is much higher than with random guessing.
Blindsight is possible due to “parallel computations” that the brain performs on visual information coming from the retina. As we remember, in the retina of the eye there is a light-receiving layer of visual receptors - rods and cones. Rods allow you to see at dusk, but provide less accurate images than cones. Cones, on the other hand, work only in fairly bright light, but they allow you to distinguish colors and finer details of a picture. Information from the rods and cones passes through several other layers of retinal cells and is processed a little in them, but this is not essential for the present story. It is important that at the exit from the retina ( lat.
retina) data paths from different rods and cones diverge. Most of it falls into the so-called retino-geniculo-striatal pathway, RGS (Fig. 1). The lion's share of the optic nerve fibers goes to the lateral geniculate body of the thalamus (LCT, or LGN in English). In Latin, knee is geniculum, hence the root “geniculo” in the name of the pathway. From the lateral geniculate bodies, signals enter the occipital regions of the cerebral cortex, the primary visual cortex (V1).
The primary visual cortex (or area 17 in Brodmann's classification) appears striped in sections under a light microscope (Fig. 2). The Latin word for stripe is stria, which is why the primary visual cortex is often called striatal ( lat.
striate cortex). (The striate cortex should not be confused with the striatum: the latter is located under the cerebral cortex and performs a completely different role - it controls movements.) In the primary visual cortex, signals from the retina are processed: neurons identify lines in the image, determine the angle of inclination of these lines and some other simple parameters Pictures. Further information goes to the secondary visual cortex (fields 18 and 19), whose neurons “catch” more complex features. Finally, the data reaches the parietal or temporal lobes. The associative areas of the cortex are located there, where full-fledged images of objects are already formed.
Rice. 2.
Primary visual (striate) cortex in sections under a light microscope. The somas (bodies) of neurons are colored with purple dye. In layer 4, the darker stria of Gennari is clearly visible - a cluster of myelinated axons - due to which the primary visual cortex received its second name - striatal. Image from Encyclopedia of Neuroscience, chapter Striate Cortex Functions
This is the fate of almost all the information that the retina brings to us. But there is an alternative way. It is called extragenicular because it does not pass through the lateral geniculate body of the thalamus (Fig. 1). Some of the fibers of the optic nerve end in the superior colliculus, or superior colliculus (SC), quadrigeminal midbrain - structures that also specialize in processing visual information. The inferior colliculi are responsible for processing sound stimuli.
Analysis of data from the retina in the superior colliculus occurs quickly, but unconsciously. Their work allows you to escape from approaching danger even before the body realizes that something is threatening it. (An amusing example of a defensive reaction provided by the superior colliculi was demonstrated by former US President George W. Bush, when at a press conference an Iraqi journalist suddenly began throwing shoes at him.) From the superior colliculus, information goes to the pillows (paired nuclei of the thalamus, see Pulvinar nuclei) , and from there - to the cerebral cortex, but not to the primary, but to the secondary (Fig. 3).
Rice.
3. Diagram of the two main pathways for transmitting visual information in the brain
It was the extragenicular visual pathway that was preserved in an unusual patient with whom Dutch and Swiss scientists worked who studied the phenomenon of blindsight. A 56-year-old man lost complete occipital cortex in both hemispheres due to two strokes occurring 6 weeks apart. This is clearly visible on MRI images (Fig. 4).
Rice. 4.
Magnetic resonance imaging (MRI) of a subject's brain (side, back, and top views).
From the discussed article in Frontiers in Human Neuroscience
Since the subject did not have an occipital cortex, he did not have a primary visual cortex. This means that the retino-geniculo-striatal visual pathway was also disrupted. A vision test confirmed this: he could not describe either the shape, color, or size of the objects that were shown to him, or the direction of their movement. But he somehow distinguished people’s faces and could move independently without touching objects.
So why was it necessary to “test” an anonymous patient if the phenomenon of false blindness is not new for a long time? Here's the thing. After some experiments on animals (see M. Jay, D. Sparks, 1984. Auditory receptive fields in primate superior colliculus shift with changes in eye position), an assumption arose that the formation of associations between sound and visual stimuli can occur without the participation of the retino- geniculo-striatal visual pathway. In other words, it is not necessary to consciously perceive the visual image of an object in order to associate it with a sound signal. Apparently, stimuli from the eyes and ears are first encountered not in the associative areas of the cortex, but in the quadrigeminal region (see M. Meredith, B. Stein, 1986. Visual, auditory, and somatosensory convergence on cells in superior colliculus results in multisensory integration) . That is, in some cases of associative learning you can do without the neocortex altogether, and this is news.
However, until recently, experiments on teaching blindsighted people to connect a visual stimulus with a sound one were carried out only on animals, and no one knew how things were going with this in humans. And then a volunteer appeared who did not have a primary visual cortex, but did have the superior colliculus. All that remains is for him to develop an association of a specific sound, which the subject obviously hears, with a specific object, which he cannot consciously perceive.
This was done as follows (Fig. 5). The subject was seated 25 cm from a gray screen, in the center of which a red circle was present for a second. For the same second, a person heard a sound with a frequency of 500 Hertz through headphones. At the same time, the sound volume gradually increased from 35 decibels (the background noise level in the room) to 80 decibels (the volume of a powerful vacuum cleaner). This experiment was repeated 270 times (they also say “270 presentations were performed”). In 2/3 of the cases, in the middle of a second, the sound volume sharply increased (up to 80 decibels) for 10 milliseconds, and then returned to its value before the sharp jump and continued to increase smoothly. The subject's task was to predict a sharp jump in volume - that is, to press a button as quickly as possible if it seemed to him that the volume was about to jump to maximum.
Rice. 5.
Experimental design.
In each pair of images, at the top
is a change in the size of the red circle,
at the bottom
is an increase in volume.
on the right
and
left
.
The dotted line
marks the appearance of a circle of maximum size, which coincided in time with a sharp increase in sound volume to maximum.
Image from the discussed article in Frontiers in Human Neuroscience
Doing this based solely on auditory information was impossible. The clue was in the size of the red circle on the screen. During the same second it could either increase or decrease. A decrease occurred 2 times more often than an increase. In addition, at the moment of a sharp jump in volume, the circle became its maximum size for 10 milliseconds, and then, like the sound, returned to its size “before the shake.” Thus, the subject should have formed an association: since the circle is decreasing, the volume will most likely soon increase sharply. The formation of this cause-and-effect relationship in the subject’s brain would be reflected in the speed of his reaction. And so it happened. Over and over again, “seeing” the shrinking circle (hint), the person, with a similar presentation, gradually pressed the button faster and faster. If the patient “saw” an enlargement of the circle (no clue), he was in no hurry to give a signal. In such cases, his reaction time also decreased from presentation to presentation (I didn’t want to wait for the volume to jump), but not so much.
Another interesting feature: blindsighted people, in principle, do not perceive violet objects (see S. Leh et al., 2006. Absence of S-cone input in human blindsight following hemispherectomy). Information from cones, which perceive light with a short wavelength (420 nm, the wavelength for violet), does not reach the superior colliculus. So the experiment described above was repeated using purple circles. In this case, the hint did not work: the decrease in the purple circle, which indicated a likely jump in loudness, had almost no effect on the subject’s reaction speed (Fig. 6). It decreased equally with any modification of the purple circle, since the subject did not perceive this figure even “on a subconscious level.”
Rice. 6.
The graph shows how the reaction time decreased in experiments with red (experiment 1) and purple (experiment 2) circles.
It can be seen that in Experiment 2, the reaction time when presented without a hint decreases in exactly the same way as with a hint. From the discussed article in Frontiers in Human Neuroscience
So, a person without a primary visual cortex can form associations between sounds and properties of visible objects. True, these objects need to be perceived by the superior colliculus, and these structures have limited capabilities (in particular, they do not allow analyzing objects that reflect light of short wavelengths (about 420 nm), that is, violet). But the main thing is that blindsighted people retain not only some “visual” skills, but also the ability, thanks to these skills, to learn new things.
Source:
Mehrdad Seirafi, Peter De Weerd, Alan J. Pegna and Beatrice de Gelder.
Audiovisual Blindsight: Audiovisual learning in the absence of primary visual cortex // Frontiers in Human Neuroscience.
2015. V. 9. P. 686.
Svetlana Yastrebova
Functions
Simplified, the visual cortex performs three main actions with visual information:
- Reception.
- Treatment.
- Broadcast.
At the initial stage, visual information is obtained and decomposed into simple parameters - brightness, color, size, and so on. The received and initially processed data is transferred to the following zones, where they are analyzed and compared with the images in memory. In this area, visual information takes on a deciphered form, that is, objects receive specific names and mental images. After processing, it is transmitted in clear images to the associative sections of the neocortex and limbic system to generate responses to the received data.
In the decision-making process, visual information is checked by data received from other sensory zones, and if it is not reliable, then it is re-evaluated. This explains the concept of visual hallucinations, when a person sees an object but does not touch it. In this case, consciousness begins to develop a set of measures to find erroneous data.
Primary area
This structure is a direct receiver of signals from the retina. Visual information initially falls on layer IV, after which it begins to be processed bit by bit and deciphered into the simplest parameters on each layer, using oriented neurons forming columns. After processing, the information is accumulated in layer VI of the cortex, from where it is transmitted to associative areas for further analysis and processing. For example, color balls are one of a group of columns that convert received information into an accessible signal indicating the color of an object.
One of the questions that remained unanswered for a long time was the mechanism of projection of the retina to certain cortical visual centers.
Initially, most neurophysiologists adhered to Monakov’s theory, according to which this projection does not have a rigid connection with a specific part of the V1 zone. He believed that the areas were interchangeable and damage to one of them would in no way affect the overall reception of visual information. However, this theory was refuted by Wilbrand and Genschen, who, after conducting a series of studies, proved that each part of the retina is projected only onto a certain part of the cortical visual center.
For quite a long time it was believed that various parts of area V1 are connected to other parts of the brain, only by horizontal fibers. However, Luke (1949), Doty (1956), Hubel (1977) and Wiesel (1982), after conducting a series of studies, determined that nearby neurons have similar functions and are combined into vertical columns or columns. This made it possible to prove that vertical connections of neurons are more important in the process of information transfer. They have a loop-like shape and different lengths, so they can be immersed in all adjacent areas of the brain.
Along with the primary processing of visual data, an equally important function of area V1 is its transfer to associative areas. It occurs simultaneously from both hemispheres through two visual pathways - dorsal and ventral.
Using the dorsal pathway, information from area V1 initially enters area V2, from there to the dorsomedial area (V6), after which it moves to area V5 and ends in the parietal lobe of the brain. This channel transmits data that characterizes the localization of the object, spatial position, the ratio of sizes to the object and the control of eye movement, that is, it answers the question - where.
Along the ventral pathway, processed data in V1 through area V2 enters area V4, from which it is transmitted to the lower part of the temporal lobe. This channel transmits information that is related to the parameters of the subject, the definition of the object, that is, it answers the question - what. In addition, this path is strongly associated with writing new data into long-term memory, as well as using existing data.
This idea of information transfer was described by Leslie Angerleider and Mortimer Mishkin. It is based on the fact that with optical illusions, information about an object is distorted, which disappears when receiving data from other senses. This theory has not been accepted by many physiologist because they consider it too simplified. However, their opinion is not confirmed by anything and most experts take this approach as a basis.
Visual cortex
Consisting of the axons of nerve cells of the lateral geniculate body (subcortical visual centers), the optic radiation conducts visual impulses coming from the homolateral halves of the retinas of both eyes to the projection zone of the visual analyzer, located in the cortex of the occipital lobe of the cerebrum, mainly on its medial surface, in the calcarine region furrows (field 17, according to Brodmann’s architectural map).
In this case, the representation of the macula of the retina and its central fovea (the place of arrival of impulses passing earlier along the papillomacular bundle) is located in the posterior part of this field (in the area of the pole of the occipital lobe); in front of it is adjacent the projection of the paramacular zone of the homolateral halves of the retinas, and then their areas located on the periphery. It is noteworthy that the projection of the retinal spot and its central fovea occupies a disproportionately large area in the primary visual cortex, moving at the same time to the convexital surface of the occipital pole.
It is estimated that the area of projection onto the visual cortex of the central fovea of the retina is 35 times larger than the size of the projection area of the same area of the retina located at some distance from it. It can also be noted that the upper parts of the homolateral halves of the retinas (upper quadrants) are projected onto the upper lip of the calcarine sulcus, which is part of the wedge (cuneus), and the lower parts - onto the lower lip of the same sulcus, which belongs to the lingual gyrus (gyrus linqualis). The clear retinotopic order of the architectonics of cortical area 17 is the reason why it is sometimes called the “cortical retina.”
The concept of visual cortex includes
- primary visual cortex (also called striate cortex or visual area V1. It is equivalent to Brodmann area 17, or BA17.
- extrastriate visual cortex (secondary visual areas) - areas V2, V3, V4, and V5. It includes Brodmann areas 18 and 19. Secondary visual areas, also called visual association areas, lie lateral, anterior, superior and inferior to the primary visual cortex. In addition, most of these areas overlap the lateral surfaces of the occipital and parietal cortices in outward folds. Secondary visual signals are transmitted to these areas to analyze their meaning. For example, all around the primary visual cortex is Brodmann area 18, where almost all signals from the primary visual cortex are sent. Brodmann area 18 is called visual area II, or simply V2. Other, more distant secondary visual areas have specific designations V3, V4, etc. (more than a dozen areas). The significance of all these areas lies in the gradual identification and analysis of various aspects of the visual image.
The visual cortex is present in each of the cerebral hemispheres. Areas of the visual cortex of the left hemisphere receive signals from the right half of the visual field, and the right hemisphere receives signals from the left half.
Cortical area 17 (according to Brodmann) is also called the primary visual cortex or striate cortex (area striata), which is due to the structural features (“striations”) of this cortical field. If the cortex of most territories of the cerebral hemispheres consists of six layers, then in the striate cortex layer IV is divided into three sublayers by Gennari stripes, consisting of myelin fibers of optic radiation penetrating here. Some morphologists believe that a similar division of its other layers is possible.
It can be assumed that this structural feature of the striate cortex is a reflection of the special complexity of its functions. In the primary visual cortex, decoding of visual information coming here from the retinas begins. It is believed that the central fovea of the macula (macula) of the retina of each eye in the primary visual cortex has bilateral cortical representation. Nerve cells located in field 17 have connections with neurons of neighboring cortical fields (18 and 19, according to Brodmann), which carry out further analysis and synthesis of elementary visual signals entering the primary visual cortex. Physiologists call these cortical fields prestriate or secondary visual cortex. The associative cortical territories of the temporal and parietal lobes adjacent to the secondary visual cortex are also involved in processing visual information and transforming it into visual images.
The secondary visual cortex and the associated associative cortical territories perceive the visible and compare it with previously acquired information already stored in memory. This ensures recognition of visual images and their assessment, the possibility of forming ideas, which in turn can stimulate the creation of a response plan and its implementation in the form of changes in the direction of gaze, facial reactions, movements, actions of varying nature and degree of complexity.
It is now generally accepted that the axons of some neurons in the visual cortex approach the cortical and subcortical oculomotor centers. It is generally accepted that reflex oculomotor cortical centers are located in the deep layers of the visual cortex itself in the occipital lobes of the brain.
The cortical centers of voluntary gaze movements are located in the posterior frontal region (the field of both hemispheres. The main subcortical oculomotor centers are the anterior colliculus of the midbrain roof (vertical gaze center) and the pontine gaze center (horizontal gaze center).
of both hemispheres. The main subcortical oculomotor centers are the anterior colliculus of the midbrain roof (vertical gaze center) and the pontine gaze center (horizontal gaze center).
Thus, under the influence of information received by the visual cortex and coming from the retina, the possibility of both reflexive and voluntary control over the state of gaze arises. Changing its direction contributes to the consistent fixation of the gaze on individual objects located in the environment, which enriches the subject’s awareness and helps expand the volume of visual information received.
If, as is commonly believed, there are approximately 1 million ganglion cells in the retina of each eye, and each lateral geniculate body contains up to 1.5 million nerve cells, then in the primary visual cortex of one cerebral hemisphere there are already about 200 million of them. This is the ratio of the number of neurons in the subcortical and cortical visual centers creates the possibility of representing a pronounced divergence of visual impulses reaching the primary visual cortex, and the presence in it of prerequisites for an in-depth analysis of this information. Further divergence of the pathway of visual information can be considered as an argument in favor of the possible implementation by the secondary visual cortex of mainly the synthesis of diverse elements of various visual sensations coming from the primary visual cortex.
For a long time it was recognized that connections between individual areas of the visual cortex and other cortical areas are carried out only through horizontally located transcortical fibers, but today this point of view is recognized as simplified. Experimental studies by Sperry and Miner (1955) demonstrated that horizontally running cortical connections are in most cases short and therefore cannot ensure the interaction of cortical areas located at a significant distance from each other. These experimenters dissected the visual cortex of cats or inserted mica strips into it vertically, which led to the division of the visual cortex into many small islands connected to each other only through the subcortical white matter. However, after this, the experimental animals, having recovered from the operation, retained the ability to see and carried out the visual differentiation they had previously mastered.
Luke (Louchs RB, 1949), Doty et al. (Doty RW et al., 1956) and other researchers have shown that connections between individual areas of the visual cortex and the rest of the brain usually take the form of loops, plunging into the brain to different depths. In this case, individual columns of cortical nerve cells can respond to visual impulses that arise when the retina of the eyes is exposed to electromagnetic waves of different length, intensity and direction. In a similar way, cortical visual neurons associated with certain areas of the retina of the right or left eye, as well as with the peripheral oculomotor system, are combined into columns. The existence of ocular dominance columns, as well as orientation columns, has now been proven (Hubel D., 1990).
D. Hubel and T.N. Wiesel (Hubel D. and WieselT.N., 1977, 1982), using microelectrode recording of the bioelectric potentials of neurons located at different depths of the visual cortex, revealed the presence of vertical excitatory connections between them. It was found that nearby neurons in the visual cortex usually have similar functional properties and are combined into vertical columns, or pillars. These authors proved that nerve cells within a narrow vertical column respond to a stimulus of only a certain modality, while cells in neighboring columns can autonomously respond to visual signals of the same or a different modality. This led to the judgment that in the visual system, groups of cortical neurons respond mainly to information that comes to them not through horizontal, but through vertical associative connections. The existence of ocular dominance columns, as well as orientation columns, has now been proven (Hubel D., 1990).
Thus, visual impulses coming from the retina, first of all, enter the primary visual cortex, which has not only horizontal, but also especially important vertical loop-shaped connections, both with the secondary visual cortex and with subcortical centers that influence visual functions.
D. Hubel (1990) identified areas of approximately 2x2 mm in the primary visual cortex. Such a section of the cortex, which he called a module, usually represents a group of 500-1000 functional units. Each module has all the mechanisms necessary to fully process information coming from the corresponding part of the retina.
It turned out that if the part of the retina associated with the module was located in the area of the central fovea of the macula, then such a part of it per cortical module (a fragment of the primary visual cortex) is usually very small. If the studied fragment of the cortex (a module having a similar area) is associated with the periphery of the retina, then it perceives signals from a region of the retina several times larger in area. It was found that the module onto which the structures of the central fovea of the retinal spot are projected and the module associated with its periphery cannot be considered homogeneous in structure. In addition to different visual acuity, as the module moves away from the projection of the central fovea of the retina, other parameters of visual perception also change. In particular, color and binocular vision decreases. At the same time, the peripheral parts of the retina, in which there is a particularly significant predominance of rods over cones and associated cortical modules, are more sensitive to weak light sources and to moving objects in the field of view.
In the process of in-depth study of the cerebral cortex, several levels of integration of visual information in it were identified. At the same time, the special importance in the implementation of this process of associative cortical zones is recognized, in which the synthesis of information obtained through not only vision, but also other senses occurs. In addition, numerous connections of the cortex with the structures of the limbic system largely determine the influence of information coming from the periphery through analyzers of various types of modality, in particular through the visual analyzer, on the state of the emotional and vegetative spheres.
- If field 17 (according to Brodmann) provides the emergence of elementary sensations (light perception, color perception, elements that make up an object located in the observed space),
- then the analysis and synthesis of these sensations and the possibility of expanding their number and definition through reflex stimulation of the degree of accommodation of the lenses and convergence of the eyes, as well as changing the position of the gaze, depend on field 18.
- At the same time, it is recognized that field 19 of the secondary visual cortex already creates the possibilities of optical-gnostic, objective and spatial perception, and contributes to the formation of ideas.
In the process of subsequent interpretation of more complex visual representations that arise in the associative zones of the cortex, they are compared with visual impressions that existed in the past and stored in memory, as well as with information received in parallel using other sensory systems.
Irritation of the primary visual cortex (field 17) is usually accompanied by the appearance of elementary visual sensations - photopsia. Its destruction leads to changes in the visual fields (variants of incomplete or complete hemianopsia, pathological scotomas in the homolateral halves of the visual fields) and to a decrease in vision, even to blindness.
Irritation of cortical fields 18 and 19, as well as the associated association zones of the temporal and parietal lobes of the cortex, is accompanied by more complex visual sensations such as illusions or hallucinations, and their destruction can cause a violation of orientation in space and a disorder of recognition of visible objects - visual agnosia.
Manifestations of visual agnosia are especially significant with bilateral dysfunction of the 18th, 19th cortical fields and adjacent associative cortical territories, as well as with damage to interhemispheric commissural connections. The patient in such cases can see and avoid obstacles, but does not recognize what is visible and does not have the ability to adequately assess and understand the surrounding situation. He tries to understand it using his remaining hearing, tactile, temperature and other types of sensitivity.
There are several types of visual agnosia
- agnosia for things - objective,
- Letter agnosia is a special form of alexia (“pure” alexia, or alexia without aphasia),
- agnosia for colors, faces, etc.
Damage to the right occipito-parietal region often leads to the development of visual disorientation in space, to complete neglect of its left half.
Due to the complexity of the structure of the cerebral cortex and the physiological processes occurring in it, the study of its functions is fraught with special difficulties. Having dedicated his life to overcoming these difficulties, the American neurophysiologist D. Hubel admitted in 1990 that, despite some successes in the study of the transformation of visual information in the primary visual cortex, “almost nothing is known about what happens at further stages of its processing.” .
Neuropsychologists, when studying human acquisition of visual information, confirmed the somatotopic order of propagation of visual signals from the retina to the primary visual cortex. It has been established that partial damage to the visual pathways or the primary visual cortex leads to the loss of strictly defined fragments of the visual fields, which in some cases can be significant in the topical diagnosis of diseases and traumatic brain lesions.
It has also been established that irritation of the primary visual cortex is accompanied by the development of photopsia in the form of flickering luminous dots or spots in certain areas of the visual field. When areas of the secondary visual cortex or its associated association zones are damaged, disorders of the perception of visual images arise, in particular micro- or macrophotopsia, complex visual hallucinosis or hallucinations are possible, as well as visual agnosia, which is manifested by a disorder in the recognition of individual visual images, impaired ability to synthesize and generalization of the received visual information, which leads to disorientation in space and inadequacy of ideas and behavioral reactions.
It has also been established that damage to individual areas of the primary visual cortex of one hemisphere does not have any serious significance for the implementation of higher mental processes and leads only to partial impairment of the visual fields, in particular to the occurrence of pathological scotomas. However, if the functions of the associative zones of the cortex of the parieto-occipital region or the deep parts of the subdominant (usually the right) hemisphere are impaired, the patient may experience ignorance of the opposite half of space and his own body, which is usually regarded as anosognosia and autotopagnosia - variants of spatial visual agnosia.
If the pathological process affects the visual pathways on both sides above the subcortical visual centers, central blindness occurs in both eyes, in which the pupillary reflexes remain intact. In cases where, due to hemodynamic disturbances in the vessels of the vertebral-basilar basin, which is relatively rare in practice, the primary visual cortex of both hemispheres is selectively affected, it is possible to preserve tubular vision on both sides due to the fact that the area of the occipital lobe cortex, which The macula (macula) of the retina is projected and has additional blood supply through the posterior branches of the middle cerebral arteries.
Visual pathways
Visual areas V1 (right and left) transmit visual information along two primary visual pathways - dorsal and ventral.
- The dorsal pathway begins in the primary visual cortex (visual area V1), passes through visual area V2, then goes to the dorsomedial visual area (DM or V6), visual area MT (otherwise called V5), and the posterior parietal cortex. The dorsal pathway (the “where?” or “how?” channel) is associated with movement, the idea of object localization, control of eye movements (saccades), and the use of visual information to assess the reach of objects and reach visible objects with the hands).
- The ventral pathway also begins in area V1 and passes through V2, but then passes through visual area V4 to the ventral (inferior) part of the temporal lobe of the cortex. The ventral pathway (the “what?” channel) is associated with the process of shape recognition, representation of an object, and also with long-term memory.
The branching of the flow of visual information into the dorsal (“where?”) and ventral (“what?”) pathways—in other words, into the “action” and “recognition” channels—was first described by Leslie Ungerleider and Mortimer Mishkin, and this hypothesis still provokes disputes among physiologists and scientists studying the phenomenon of vision. It may oversimplify actual processes in the visual cortex. It is based on the discovery that optical illusions such as the Ebbinghaus illusion can distort correct perception, but in cases where the subject responds to a visual stimulus with an action, such as grasping the object seen, the image is perceived without distortion. However, there is a 2005 publication stating that both cortical systems for processing visual information - both the “action channel” and the “recognition channel” - are equally susceptible to illusion deception.
Neurons in the visual cortex generate action potentials when visual stimuli impinge on their receptive fields. The receptive field is defined as the area of the visual field whose stimulation results in the generation of an action potential. But sometimes a neuron can respond better to a particular set of visual stimuli presented within its receptive field. This property is called neuron selectivity. In older visual areas, neuronal selectivity is low. Thus, a neuron in visual area V1 can be excited in response to any vertical stimulus in its receptive field. Neurons of integrative visual areas are characterized by complex selectivity. For example, neurons in the visual integrative area of the superior temporal sulcus (in monkeys) or the ventral surface of the fusiform gyrus at the border between the occipital and temporal lobes (in humans) can be excited only when the receptive field is stimulated by images of faces.
The blood supply to the visual cortex is carried out mainly by the calcarine branch (lat. ramus calcarinus) of the medial occipital artery - a branch of the terminal (cortical) part (lat. pars corticalis) of the posterior cerebral artery (lat. arteria cerebri posterior). The calcarine branch lies in the calcarine groove (lat. sulcus calcarinus, fissura calcarina) of the cerebral cortex.
Secondary area
The term secondary cortex refers to the totality of all associative areas that analyze, decipher and complement the received visual information from the V1 sensory area. In addition, more distant zones are directly involved in shaping the reaction to the received data.
Processing and analyzing information in these zones is possible due to their structure, which differs from V1 in the presence of a large number of neurons that are capable of performing complex operations. It is worth noting that the further the zone is from V1, the greater the number of complex neurons it contains.
The modern understanding of this area contains the message that there are levels of information integration in it. Following this hypothesis, each zone increases and refines the data passing through it, and the greater the number of them that took part in processing, the more complex and final the images are.
In addition, recent studies have shown that a number of axons of neurons in area V2 connect to gaze control centers. This indicates that in the process of analyzing and decoding data, the secondary visual cortex has the ability to supplement it. To do this, she reflexively controls the accommodation of the lenses and the direction of gaze. Also, in addition to reflex control, more distant associative zones can purposefully regulate the position of the gaze and focus attention on the object of visualization.
Along with the enormous efforts expended on the study of associative zones, one of the most famous neurophysiologists believed D. Hubel that all science had not advanced even a couple of percent towards the study of the processes of analysis and processing in the secondary structure of the cortex.