Зрение – это одно из основных чувств человека, которое влияет практически на все аспекты жизни: ориентация в пространстве, получение информации об окружающих предметах, обучение и развлечение, распознавание эмоций других людей и многое другое.
Роль управляющего центра данного чувства выполняет зрительная кора головного мозга. Множество заболеваний зрения происходят по причине нарушений в работе данной области. Поэтому для нахождения методик лечения данных болезней большое внимание уделяется изучению строения и выполняемых функций зрительной коры .
Общие сведения
Зрительная кора – участок коры головного мозга, который принимает, анализирует и обрабатывает визуальную информацию об окружающем мире. Она тесно связана с ассоциативными областями, которые отвечают за высшие нервные функции.
Расположен этот участок в затылочной и срединой области обоих полушарий и занимает 17-ое, 18-ое и 19-ое цитоархитектонические поля Бродмана.
Визуальные данные с левой половины зрительного потока принимается участками коры правого полушария, а правого – левыми.
Органы чувств (анализаторы). Строение и функции органов зрения и слуха
Органы чувств (анализаторы). Строение и функции органов зрения и слуха
Анализатор
— функциональная система, состоящая из:
— рецептора,
— чувствительного проводящего пути
— соответствующей зоны коры, куда проецируется данный вид чувствительности.
Анализ и синтез полученной информации осуществляются в строго определенном участке — зоне коры больших полушарий
.
По особенностям клеточного состава и строения кору больших полушарий разделяют на ряд участков, называемых корковыми полями
. Функции отдельных участков коры неодинаковы. Каждому рецепторному аппарату на периферии соответствует область в коре —
корковое ядро анализатора.
Важнейшие зоны коры следующие:
• Двигательная зона
расположена в переднецентральной и заднецентральной областях коры (передней центральной извилине впереди центральной борозды лобной доли).
• Чувствительная зона
(зона кожно-мышечной чувствительности расположена позади центральной борозды, в задней центральной извилине теменной доли). Наибольшую площадь занимает корковое представительство рецепторов кисти и большого пальца руки, голосового аппарата и лица, наименьшую — представительство туловища, бедра и голени.
• Зрительная зона
сосредоточена в затылочной доле коры. В нее поступают импульсы от сетчатки глаза, она осуществляет различение зрительных раздражений.
• Слуховая зона
расположена в верхней височной извилине височной доли.
• Обонятельная и вкусовая зоны
— в переднем отделе (на внутренней поверхности) височной доли каждого полушария.
Деятельность коры головного мозга человека и высших животных определена И.П. Павловым как высшая нервная деятельность
, представляющая собой условно-рефлекторную функцию коры головного мозга.
Анализаторы
– совокупность нервных образований, обеспечивающих осознание и оценку, действующих на организм, раздражителей. Анализатор состоит из воспринимающих раздражение рецепторов, проводящей части и центральной части – определенной области коры головного мозга, где формируются ощущения.
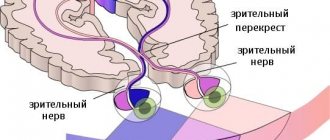
Зрительный анализатор
обеспечивает получение зрительной информации из окружающей среды и состоит из трех частей:
периферической – глаз, проводниковой – зрительного нерва центральной – подкорковой и зрительной зоны коры головного мозга.
Глаз
состоит из глазного яблока и вспомогательного аппарата, к которому относятся веки, ресницы, слезные железы и мышцы глазного яблока.
Глазное яблоко
расположено в глазнице и имеет шаровидную форму и 3
оболочки
:
фиброзную,
задний отдел которой образован непрозрачной
белочной
оболочкой (
склерой
),
сосудистую,сетчатую
Часть сосудистой оболочки, снабженная пигментами, называется радужной оболочкой
.В центре радужной оболочки находится
зрачок
, который может изменять диаметр своего отверстия за счет сокращения глазных мышц.
Задняя часть сетчатки
воспринимает световые раздражения.
Передняя ее часть
– слепая и не содержит светочувствительных элементов.
Светочувствительными элементами
сетчатки являются:
палочки
(обеспечивают зрение в сумерках и темноте),
колбочки
(рецепторы цветового зрения, работающие при высокой освещенности). Колбочки расположены ближе к центру сетчатки (желтое пятно), а палочки концентрируются на ее периферии. Место выхода зрительного нерва называется
слепым пятном
. Полость глазного яблока заполнена
стекловидным телом
.
Хрусталик
имеет форму двояковыпуклой линзы. Он способен изменять свою кривизну при сокращениях ресничной мышцы. При рассматривании близких предметов хрусталик сжимается, при рассматривании отдаленных – расширяется. Такая способность хрусталика называется
аккомодацией
. Между роговицей и радужкой находится
передняя камера глаза
, между радужкой и хрусталиком –
задняя камера
. Обе камеры заполнены прозрачной жидкостью. Лучи света, отражаясь от предметов, проходят через роговицу, влажные камеры, хрусталик, стекловидное тело и, благодаря преломлению в хрусталике, попадают на
желтое пятно
сетчатки – место наилучшего видения. При этом возникает
действительное, обратное, уменьшенное изображение предмета
.
От сетчатки по зрительному нерву импульсы поступают в центральную часть анализатора – зрительную зону коры мозга
, расположенную в затылочной доле. В коре информация, полученная от рецепторов сетчатки, перерабатывается и человек воспринимает естественное отражение объекта.
Нормальное зрительное восприятие
обусловлено:
– достаточным световым потоком;
– фокусированием изображения на сетчатке (фокусирование перед сетчаткой означает близорукость, а за сетчаткой – дальнозоркость);
– осуществлением аккомодационного рефлекса.
Важнейшим показателем зрения
является его острота, т.е. предельная способность глаза различать мелкие объекты.
Аккомодация —
приспособление глаза к видению различно удаленных предметов. При аккомодации сокращаются мышцы, которые изменяют кривизну хрусталика. При постоянной избыточной кривизне хрусталика световые лучи преломляются перед сетчаткой и в результате возникает
близорукость.
Если же кривизна хрусталика недостаточна, то световые лучи фокусируются за сетчаткой и возникает
дальнозоркость.
Близорукость развивается при увеличенной продольной оси глаза. Параллельные лучи, идущие от далеких предметов, собираются (фокусируются) впереди сетчатки, на которую попадают расходящиеся лучи и в результате получается расплывчатое изображение. При близорукости назначают очки с рассеивающими двояковогнутыми стеклами, уменьшающими преломление лучей настолько, что изображение предметов возникает на сетчатке. Дальнозоркость наблюдается при укороченной оси глазного яблока. Изображение фокусируется позади сетчатки. Для исправления зрения требуются двояковыпуклые стекла. Старческая дальнозоркость развивается обычно после 40 лет, когда хрусталик теряет эластичность, твердеет и утрачивает способность менять кривизну, что мешает четко видеть на близком расстоянии. Глаз утрачивает способность к ясному видению разноудаленных предметов.
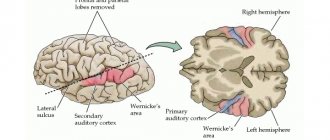
Орган слуха и равновесия.
Слуховой анализатор
обеспечивает восприятие звуковой информации и ее обработку в центральных отделах коры головного мозга.
Периферическую часть
анализатора образуют: внутренне ухо и слуховой нерв.
Центральная часть
образована подкорковыми центрами среднего и промежуточного мозга и височной зоной коры.
Ухо
– парный орган, состоящий из:
Наружного уха
– включает ушную раковину, наружный слуховой проход и барабанную перепонку.
Среднего уха
– состоит из барабанной полости, цепочки слуховых косточек и слуховой (евстахиевой) трубы. Слуховая труба связывает барабанную полость с полостью носоглотки. Это обеспечивает выравнивание давления по обеим сторонам барабанной перепонки. Слуховые косточки – молоточек, наковальня и стремечко связывают барабанную перепонку с перепонкой овального окна, ведущего в улитку. Среднее ухо обеспечивает передачу звуковых волн из среды с низкой плотностью (воздух) в среду с высокой плотностью (эндолимфу), в которой находятся рецепторные клетки внутреннего уха.
Внутреннего уха
– расположено в толще височной кости и состоит из костного и расположенного в нем перепончатого лабиринта. Пространство между ними заполнено перилимфой, а полость перепончатого лабиринта – эндолимфой. В костном лабиринте различают три отдела –
преддверие, улитку и полукружные каналы
. К органу слуха относится
улитка
– спиральный канал в 2,5 оборота. Полость улитки разделена перепончатой основной мембраной, состоящей из волоконец разной длины. На основной мембране находятся рецепторные
волосковые клетки
. Колебания барабанной перепонки передаются слуховым косточкам. Они усиливают эти колебания почти в 50 раз и через овальное окошко передаются в жидкость улитки, где воспринимаются волоконцами основной мембраны. Рецепторные клетки улитки воспринимают раздражение, поступающее от волоконец и по слуховому нерву передают его в височную зону коры головного мозга. Ухо человека воспринимает звуки частотой от 16 до 20 000 Гц.
Орган равновесия
или
вестибулярный аппарат
образован двумя
мешочками
, заполненными жидкостью, и
тремя полукружными каналами
. Рецепторные
волосковые клетки
расположены на дне и внутренней стороне мешочков. К ним примыкает мембрана с кристаллами – отолитами, содержащими ионы кальция. Полукружные каналы расположены в трех взаимно перпендикулярных плоскостях. В основаниях каналов находятся волосковые клетки. Рецепторы отолитового аппарата реагируют на ускорение или замедление прямолинейного движения. Рецепторы полукружных каналов раздражаются при изменениях вращательных движений. Импульсы от вестибулярного аппарата по вестибулярному нерву поступают в ЦНС. Сюда же поступают импульсы от рецепторов мышц, сухожилий, подошв. Функционально вестибулярный аппарат связан с мозжечком, отвечающим за координацию движений, ориентацию человека в пространстве.
Вкусовой анализатор
состоит из рецепторов, расположенных во вкусовых почках языка, нерва, проводящего импульс в центральный отдел анализатора, который находится на внутренних поверхностях височной и лобной долей.
Обонятельный анализатор
представлен обонятельными рецепторами, находящимися в слизистой оболочке носа. По обонятельному нерву сигнал от рецепторов поступает в обонятельную зону коры головного мозга, находящуюся рядом со вкусовой зоной.
Кожный анализатор
состоит из рецепторов, воспринимающих давление, боль, температуру, прикосновение, проводящих путей и зоны кожной чувствительности, расположенной в задней центральной извилине.
Анатомия структуры зрения
Как и все остальные области головного мозга, отвечающие за органы чувств, эта область разделяется на сенсорные участки и ассоциативные. Сенсорная область имеет название первичная зрительная кора (стриарная или зона V1), а ассоциативная – вторичная зрительная кора (престриатная).
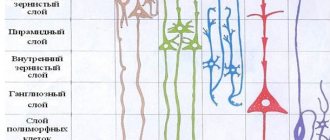
Зона V1 находиться в районе шпорной борозды, и растянута вдоль медиальной стороны. Ее положение соответствует 17-ому полю Бродмана. Как и практически весь неокортекс, она состоит из 6 горизонтальных слоев, однако в IV слое имеются существенные отличия от остальных областей. IV слой в данном случае разделен на четыре подслоя. Деление происходит полосками Дженари, которые состоят из миелиновых волокон. Кроме того, существует теория о разделении и остальных слоев на большее количество подслоев, однако на сегодняшний день, данная гипотеза не подтверждена.
Престриатная кора состоит из небольших ассоциативных областей, которые лежат в стороне от центральной плоскости и опоясывают зону V1. На сегодняшний момент выделяют около 30 различных ассоциативных участков, которые обозначаются латинской буквой V и цифрой после ее, указывающей отдалённость от первичной. Первый участок, который вплотную лежит к зоне V1 имеет обозначение V2. Все зрительные ассоциативные области соответствуют 18-ому и 19-ому полям Бродмана.
Строение областей престриатной коры практически идентично всему неокортексу. Они также имеют 6 горизонтальных слоев и ряды вертикальных колонок. Единственным отличием является, то, что у данных областей, VI слой имеет более насыщенную и сложную архитектуру.
Человек без зрительной коры способен связать звук и зрительный стимул
Рис. 1.
Пути передачи зрительной информации в мозге. Th (thalamus) — таламус; V1 (visual 1) — первичная зрительная кора; SC (superior colliculus) — один из двух верхних бугорков четверохолмия; Pulv (pulvinar) — подушка; LGN (lateral geniculate nucleus) — латеральное коленчатое тело. Также на рисунке показаны вентральный и дорсальный пути передачи информации в коре; о них более подробно рассказано в новости Мозг сортирует зрительные образы не так, как считалось ранее («Элементы», 23.08.2013). Изображение из статьи N. Diederich et al., 2014. Are patients with Parkinson’s disease blind to blindsight?
Кора больших полушарий (неокортекс) нужна нам для того, чтобы устанавливать связи между информацией, поступающей через органы чувств. В неокортексе складываются образы предметов (например, банан не только жёлтый, но и гладкий), и именно благодаря ему собака Павлова смогла понять, что еда появляется не сама по себе, а только вслед за включением лампочки. Однако новые исследования неврологических пациентов показывают, что ассоциативное обучение возможно и без участия неокортекса. Так, человек, который лишился зрительной коры, может установить ассоциацию между кругом определённого цвета и размера и звуком определённой высоты и громкости — и это при том, что сознательно он не воспринимает цвета, размеры и формы предметов.
Феномен псевдослепоты, или слепозрения (см. Blindsight), известен уже несколько десятков лет. А в последние годы его популяризации немало способствовал одноимённый научно-фантастический роман Питера Уоттса, специалиста по биологии моря (см. “Blindsight”, «Ложная слепота») У слепозрячих всё в порядке с глазами, но нарушена работа первичной зрительной коры больших полушарий. Поэтому такие люди не различают цвета и формы объектов, а также не понимают, движутся эти объекты или нет. Тем не менее слепозрячие способны самостоятельно передвигаться, обходя препятствия, и могут описать предмет, стоящий перед ними. И хотя испытуемые утверждают, что не видят никакого предмета, а только угадывают его характеристики, процент верных ответов получается гораздо выше, чем при случайном угадывании.
Слепозрение возможно благодаря «параллельным вычислениям», которые мозг производит над зрительной информацией, поступающей из сетчатки. Как мы помним, в сетчатке глаза есть световоспринимающий слой из зрительных рецепторов — палочек и колбочек. Палочки позволяют видеть в сумерках, но дают меньшую точность изображения, чем колбочки. Колбочки, наоборот, работают только при достаточно ярком освещении, но позволяют различать цвета и более мелкие детали картинки. Информация от палочек и колбочек проходит через несколько других слоёв клеток сетчатки и немного обрабатывается в них, но для настоящего повествования это не существенно. Важно, что на выходе из сетчатки (лат.
retina) пути данных от разных палочек и колбочек расходятся. Большая часть попадает в так называемый ретино-геникуло-стриарный путь, РГС (рис. 1). Львиная доля волокон зрительного нерва идёт в латеральные коленчатые тела таламуса (ЛКТ, или LGN по-английски). По-латыни колено — geniculum, отсюда корень «геникуло» в названии проводящего пути. Из латеральных коленчатых тел сигналы поступают в затылочные области коры больших полушарий, в первичную зрительную кору (V1).
Первичная зрительная кора (или поле 17 в классификации Бродмана) на срезах под световым микроскопом выглядит полосатой (рис. 2). Полоска по-латыни — stria, поэтому и первичную зрительную кору часто называют стриарной (лат.
striate cortex). (Стриарную кору не стоит путать со стриатумом: последний находится под корой больших полушарий и выполняет совершенно другую роль — контролирует движения.) В первичной зрительной коре сигналы от сетчатки обрабатываются: нейроны выделяют в изображении линии, определяют угол наклона этих линий и некоторые другие простые параметры картинки. Дальше информация идёт во вторичную зрительную кору (поля 18 и 19), нейроны которой «вылавливают» более сложные признаки. Наконец, данные попадают в теменные или височные доли. Там расположены ассоциативные области коры, где уже складываются полноценные образы предметов.
Рис. 2.
Первичная зрительная (стриарная) кора на срезах под световым микроскопом. Фиолетовым красителем окрашены сомы (тела) нейронов. В слое 4 хорошо видна более темная полоска Дженнари (Stria of Gennari) — скопление миелиновых аксонов, — благодаря которой первичная зрительная кора получила свое второе название — полосатая (стриарная). Изображение из книги Encyclopedia of Neuroscience, глава Striate Cortex Functions
Такова судьба почти всей информации, которую приносит нам сетчатка. Но есть и альтернативный путь. Его называют экстрагеникулярным, так как он не проходит через латеральные коленчатые тела таламуса (рис. 1). Часть волокон зрительного нерва заканчивается в верхних бугорках, или в верхнем двухолмии (Superior colliculus, SC), четверохолмия среднего мозга — структурах, которые тоже специализируются на обработке зрительной информации. Нижние бугорки четверохолмия отвечают за обработку звуковых стимулов.
Анализ данных от сетчатки в верхних бугорках идёт быстро, но бессознательно. Их работа позволяет уйти от приближающейся опасности ещё до того, как организм понял, что ему что-то угрожает. (Забавный пример защитной реакции, обеспеченной верхними бугорками, продемонстрировал бывший президент США Джордж Буш, когда на пресс-конференции иракский журналист внезапно начал кидать в него ботинки.) Из верхних бугорков четверохолмия информация идёт в подушки (парные ядра таламуса, см. Pulvinar nuclei), а уже оттуда — в кору больших полушарий, но не в первичную, а во вторичную (рис. 3).
Рис. 3.
Схема двух основных путей передачи зрительной информации в мозге
Именно экстрагеникулярный зрительный путь сохранился у необычного пациента, с которым работали голландские и швейцарские учёные, исследовавшие феномен слепозрения. Мужчина 56 лет полностью потерял затылочную кору в обоих полушариях из-за двух инсультов, произошедших с интервалом в 6 недель. Это хорошо видно на снимках МРТ (рис. 4).
Рис. 4.
Изображение мозга испытуемого, полученное с помощью магнитно-резонансной томографии (вид сбоку, сзади и сверху). Из обсуждаемой статьи в
Frontiers in Human Neuroscience
Поскольку у испытуемого не было затылочной коры, у него не было и первичной зрительной коры. А значит, был нарушен и ретино-геникуло-стриарный зрительный путь. Проверка зрения подтверждала это: он не мог описать ни форму, ни цвет, ни размер предметов, которые ему показывали, ни направление их движения. Зато он каким-то образом различал лица людей и мог самостоятельно передвигаться, не трогая предметы.
Так для чего же было «испытывать» анонимного пациента, если феномен ложной слепоты давно не новость? А дело вот в чём. После некоторых экспериментов на животных (см. M. Jay, D. Sparks, 1984. Auditory receptive fields in primate superior colliculus shift with changes in eye position) возникло предположение, что формирование ассоциаций между звуковыми и зрительными стимулами может происходить и без участия ретино-геникуло-стриарного зрительного пути. Иными словами, не обязательно сознательно воспринимать зрительный образ предмета, чтобы проассоциировать его со звуковым сигналом. Судя по всему, стимулы от глаз и ушей впервые встречаются не в ассоциативных областях коры, а в четверохолмии (см. M. Meredith, B. Stein, 1986. Visual, auditory, and somatosensory convergence on cells in superior colliculus results in multisensory integration). То есть в некоторых случаях ассоциативного обучения можно и вовсе обойтись без неокортекса, а это новость.
Впрочем, до недавних пор эксперименты по обучению слепозрячих связывать зрительный стимул со звуковым велись только на животных, а как c этим обстоят дела у человека — никто не знал. И вот появился доброволец, у которого первичной зрительной коры нет, но есть верхние бугорки четверохолмия. Осталось только выработать у него ассоциацию конкретного звука, который испытуемый заведомо слышит, с конкретным предметом, который он сознательно воспринять не может.
Делали это следующим образом (рис. 5). Испытуемого сажали в 25 см от серого экрана, в центре которого в течение секунды присутствовал красный кружок. На протяжении той же самой секунды человек слышал через наушники звук частотой 500 Герц. При этом громкость звука равномерно увеличивалась от 35 децибел (фоновый уровень шума в комнате) до 80 децибел (громкость мощного пылесоса). Такой эксперимент повторяли 270 раз (ещё говорят «провели 270 предъявлений»). В 2/3 случаев в середине секунды громкость звука резко увеличивалась (до 80 децибел) на 10 миллисекунд, а потом возвращалась к своему значению до резкого скачка и продолжала плавно нарастать. Задача испытуемого заключалась в том, чтобы предугадать резкий скачок громкости — то есть как можно быстрее нажать на кнопку, если ему казалось, что громкость вот-вот подскочит до максимума.
Рис. 5.
Схема эксперимента. В каждой паре изображений
вверху
— изменение размера красного круга,
внизу
— увеличение громкости.
Справа
и
слева
указано число предъявлений одного типа.
Пунктиром
отмечено появление круга максимального размера, которое по времени совпадало с резким увеличением громкости звука до максимума. Изображение из обсуждаемой статьи в
Frontiers in Human Neuroscience
Сделать это, руководствуясь только слуховой информацией, было невозможно. Подсказка крылась в размерах красного кружка на экране. Он на протяжении той же самой секунды мог либо увеличиться, либо уменьшиться. Уменьшение происходило в 2 раза чаще, чем увеличение. Кроме того, в момент резкого скачка громкости кружок на 10 миллисекунд становился максимального размера, а затем, как и звук, возвращался к своей величине «до встряски». Таким образом, у испытуемого должна была возникнуть ассоциация: раз круг уменьшается, громкость, скорее всего, скоро резко прибавится. Формирование этой причинно-следственной связи в мозге испытуемого отразилось бы на скорости его реакции. Так и произошло. Раз за разом «видя» уменьшение круга (подсказку), человек при аналогичном предъявлении постепенно быстрее и быстрее нажимал на кнопку. Если же пациент «видел» увеличение круга (отсутствие подсказки), он не торопился подавать сигнал. В таких случаях время его реакции тоже снижалось от предъявления к предъявлению (не хотелось дожидаться скачка громкости), но не так сильно.
Ещё одна интересная особенность: слепозрячие в принципе не воспринимают фиолетовые предметы (см. S. Leh et al., 2006. Absence of S-cone input in human blindsight following hemispherectomy). Информация от колбочек, воспринимающих свет с короткой длиной волны (420 нм, длина волны для фиолетового), не приходит в верхние бугорки четверохолмия. Так что эксперимент, описанный выше, повторили, используя фиолетовые кружки. В данном случае подсказка не сработала: уменьшение фиолетового кружка, которое указывало на вероятный скачок громкости, почти не отражалось на скорости реакции испытуемого (рис. 6). Она одинаково уменьшалась при любых видоизменениях фиолетового кружка, так как эту фигуру испытуемый не воспринимал даже «на подсознательном уровне».
Рис. 6.
График показывает, как снижалось время реакции в опытах с красными (эксперимент 1) и фиолетовыми (эксперимент 2) кружками. Видно, что в эксперименте 2 время реакции при предъявлении без подсказки снижается точно так же, как и с подсказкой. Из обсуждаемой статьи в
Frontiers in Human Neuroscience
Итак, человек без первичной зрительной коры может формировать ассоциации между звуками и свойствами видимых объектов. Правда, нужно, чтобы эти объекты воспринимались верхними бугорками четверохолмия, а эти структуры имеют ограниченные возможности (в частности, они не позволяют анализировать объекты, отражающие свет коротких длин волн (около 420 нм), то есть фиолетовые). Но главное, что у слепозрячих сохраняются не только некоторые «зрительные» навыки, но и способность благодаря этим навыкам обучаться новому.
Источник:
Mehrdad Seirafi, Peter De Weerd, Alan J. Pegna and Beatrice de Gelder. Audiovisual Blindsight: Audiovisual learning in the absence of primary visual cortex //
Frontiers in Human Neuroscience.
2015. V. 9. P. 686.
Светлана Ястребова
Функции
Упрощенно зрительная кора выполняет три основных действия с визуальной информацией:
- Прием.
- Обработка.
- Передача.
На начальном этапе визуальная информация получается и раскладывается на простые параметры – яркость, цвет, размер и так далее. Полученные и первично обработанные данные передаются в следующие зоны, в которых происходит их анализ и сравнение с имеющимися в памяти образами. На этом участке визуальная информация приобретает расшифрованный вид, то есть предметы получают конкретные названия и мысленные образы. После обработки, она понятными образами передается в ассоциативные отделы неокортекса и лимбическую систему для генерации ответных действий на полученные данные.
В процессе принятия решений, визуальная информация проверяется данными поступившими от других сенсорных зон и если она не достоверна, то происходит ее переоценка. Это объясняет понятие визуальных галлюцинаций, когда человек видит предмет, но не осязает его. В этом случае сознание начинает вырабатывать комплекс мер для нахождения ошибочных данных.
Первичная область
Данная структура является непосредственным приемником сигналов с сетчатки глаза. Визуальная информация изначально попадает на IV слой , после чего она начинает по битам обрабатываться и расшифровывается на простейшие параметры на каждом слое, при помощи ориентированных нейронов образующих колонки. После обработки сведениия накапливаются в VI слое коры, откуда передаются на ассоциативные области для дальнейшего анализа и обработки. Например, цветовые шарики – одна из групп колонок, которые преобразуют полученную информацию в доступный сигнал указывающий цвет предмета.
Одним из вопросов, на который долгое время не было ответа, являлся механизм проекции сетчатки глаза на определенные кортикальные зрительные центры.
Изначально большинство нейрофизиологов придерживалось теории Монакова, по которой данная проекция не имеет жесткую привязку с определенным участком зоны V1. Он считал, что участки взаимозаменяемые и повреждение одного из них, ни в коем случае не повлияет на общий прием зрительной информации. Однако данная теория была опровергнута Вильбрандом и Геншеном, которые, проведя ряд исследований, доказали, что каждый участок сетчатки проецируется только на определенный участок кортикального зрительного центра.
Достаточно долго считалось, что различные участки зоны V1 связаны с другими участками головного мозга, лишь горизонтальными волокнами. Однако Луке(1949г.), Доти (1956г.), Хьюбел (1977г.) и Визел (1982г.) проведя ряд исследований, определили, что рядом расположенные нейроны имеют схожие функции и объединяются в вертикальные столбы или колонки. Это дало возможность доказать, что более важными в процессе передачи информации являются вертикальные связи нейронов. Они имеют петлеобразную форму и различную длину, благодаря чему могут погружаться во все прилегающие области головного мозга.
Наряду с первичной обработкой визуальных данных, не менее важной функцией зоны V1, является передача их в ассоциативные области. Она происходит одновременно с двух полушарий двумя зрительными путями – дорсальным и вентральным.
При помощи дорсального пути информация с области V1 изначально поступает в зону V2, оттуда в дорсомедиальную зону (V6), после чего переходит в зону V5 и заканчивает путь в теменной доле головного мозга. По данному каналу передаются данные, которые характеризуют локализацию предмета, пространственного положения, соотношение размеров до объекта и управление движения глаз, то есть отвечает на вопрос – где.
По вентральному пути обработанные данные в V1 через зону V2 поступают в зону V4, из которой передаются в нижнюю часть височной доли. Данный канал передает информацию, которая связана с параметрами предмета, определения объекта, то есть отвечает на вопрос – что. Кроме того данный путь сильно связан с записью в долгосрочную память новых данных, а также использования уже имеющихся в ней.
Данное представление о передачи информации было описано Лесли Анджерлейдером и Мортимером Мишкиным. Оно основывается на том, что при оптических обманах зрения происходит искажение информации о предмете, которое пропадает при получении данных из других органов чувств. Данная теория не была принята многими физиология из-за того, что они считают ее слишком упрощённой. Однако, их мнение ничем не подтверждены и большинство специалистов, за основу принимает именно такой подход.
Зрительная кора
Состоящая из аксонов нервных клеток латерального коленчатого тела (подкорковых зрительных центров) зрительная лучистость проводит зрительные импульсы, идущие от гомолатеральных половин сетчаток обоих глаз к проекционной зоне зрительного анализатора, расположенной в коре затылочной доли большого мозга, главным образом на медиальной ее поверхности, в области шпорной борозды (поле 17, по архитектонической карте Бродманна).
При этом представительство желтого пятна сетчатки и его центральной ямки (место поступления импульсов, проходящих ранее по папилломакулярному пучку) расположено в задней части этого поля (в районе полюса затылочной доли); спереди к ней примыкает проекция парамакулярной зоны гомолатеральных половин сетчаток, а затем и их участков, расположенных на периферии. Обращает на себя внимание, что проекция пятна сетчатки и его центральной ямки занимает в первичной зрительной коре непропорционально большую площадь, переходя при этом на конвекситальную поверхность затылочного полюса.
Подсчитано, что область проекции на зрительную кору центральной ямки сетчатки в 35 раз превышает размеры зоны проекции такого же по площади участка сетчатки, находящегося на некотором расстоянии от нее. Можно отметить также, что верхние части гомолатеральных половин сетчаток (верхние квадранты) проецируются на верхнюю губу шпорной борозды, входящую в состав клина (cuneus), а нижние — на нижнюю губу той же борозды, относящуюся к язычной извилине (gyrus linqualis). Четкий ретинотопический порядок архитектоники коркового поля 17 является поводом к тому, что его иногда называют «корковой сетчаткой».
Понятие зрительная кора включает
- первичную зрительную кору(также называемую стриарной корой или зрительной зоной V1. Она эквивалентна полю Бродмана 17, или BA17.
- экстрастриарную зрительную кору (вторичные зрительные области) — зоны V2, V3, V4, и V5. Она включает поля Бродмана 18 и 19. Вторичные зрительные области, называемые также зрительными ассоциативными зонами, лежат латерально, впереди, выше и ниже по отношению к первичной зрительной коре. Кроме того, большая часть этих областей накладывается на латеральные поверхности затылочной и теменной коры в виде складок, направленных наружу. В эти области проводятся вторичные зрительные сигналы для анализа их значения. Например, со всех сторон вокруг первичной зрительной коры находится поле Бродмана 18, куда направляются практически все сигналы от первичной зрительной коры. Поле Бродмана 18 называют зрительной областью II, или просто V2. Другие, более отдаленные вторичные зрительные области имеют специфические обозначения V3, V4 и т.д. (более дюжины областей). Значение всех этих областей заключается в постепенном выявлении и анализе различных аспектов зрительного образа.
Зрительная кора присутствует в каждом из полушарий головного мозга. Области зрительной коры левого полушария получают сигналы от правой половины зрительного поля, правого полушария — от левой половины.
Корковое поле 17 (по Бродманну) именуется также первичной зрительной корой или стриарной корой (area striata), что обусловлено особенностями строения («полосатостью») этого коркового поля. Если кора большинства территорий больших полушарий состоит из шести слоев, то в стриарной коре IV слой ее разделен на три подслоя полосками Дженнари, состоящими из проникающих сюда миелиновых волокон зрительной лучистости. Некоторые морфологи считают возможным подобное разделение и других ее слоев.
Можно предполагать, что такая особенность строения стриарной коры является отражением особой сложности ее функций. В первичной зрительной коре начинается расшифровка поступающей сюда из сетчаток зрительной информации. Есть мнение, что центральная ямка пятна (желтого пятна) сетчатки каждого глаза в первичной зрительной коре имеет двустороннее корковое представительство. Нервные клетки, расположенные в поле 17, имеют связи с нейронами соседних корковых полей (18 и 19, по Бродманну), осуществляющими дальнейший анализ и синтез поступающих в первичную зрительную кору элементарных зрительных сигналов. Физиологи называют эти корковые поля престриарной или вторичной зрительной корой. Прилежащие к вторичной зрительной коре ассоциативные корковые территории височной и теменной долей также участвуют в переработке зрительной информации и в трансформации ее в зрительные образы.
Вторичная зрительная кора и прилежащие к ней ассоциативные корковые территории осуществляют восприятие видимого и сопоставление его с уже хранящейся в памяти ранее приобретенной информации. Это обеспечивает узнавание зрительных образов и их оценку, возможность формирования представлений, которые в свою очередь могут стимулировать создание плана ответных реакций и его реализацию в форме изменений направленности взора, мимических реакций, движений, действий, имеющих различный характер и степень сложности.
В настоящее время общепризнано также, что аксоны части нейронов зрительной коры подходят к корковым и подкорковым глазодвигательным центрам. При этом принято считать, что рефлекторные глазодвигательные корковые центры находятся и в глубоких слоях самой зрительной коры в затылочных долях мозга.
Корковые центры произвольных движений взора располагаются в заднелобной области (поле  обоих полушарий. Основными подкорковыми глазодвигательными центрами признают переднее двухолмие крыши среднего мозга (центр вертикального взора) и мостовой центр взора (центр горизонтального взора).
обоих полушарий. Основными подкорковыми глазодвигательными центрами признают переднее двухолмие крыши среднего мозга (центр вертикального взора) и мостовой центр взора (центр горизонтального взора).
Таким образом, под влиянием получаемой зрительной корой информации, поступающей из сетчатки, возникает возможность как рефлекторного, так и произвольного контроля за состоянием взора. Изменение его направленности способствует последовательной фиксации взора на отдельных объектах, расположенных в окружающей среде, что обогащает информированность субъекта и способствует расширению объема получаемой зрительной информации.
Если, как принято считать, ганглиозных клеток в сетчатке каждого глаза приблизительно 1 млн, а в каждом латеральном коленчатом теле содержится до 1,5 млн нервных клеток, то в первичной зрительной коре одного большого полушария мозга их уже около 200 млн. Такое соотношение числа нейронов в подкорковом и корковом зрительных центрах создает возможность представления о выраженной дивергенции зрительных импульсов, достигающих первичной зрительной коры, и наличие в ней предпосылок к углубленному анализу этой информации. Дальнейшая дивергенция пути проведения зрительной информации может рассматриваться как аргумент в пользу возможного осуществления вторичной зрительной корой главным образом синтеза поступающих из первичной зрительной коры многообразных элементов различных зрительных ощущений.
Длительное время признавалось, что связи между отдельными участками зрительной коры и другими корковыми зонами осуществляются только за счет горизонтально расположенных транскортикальных волокон, однако на сегодня такая точка зрения признается упрощенной. Экспериментальные исследования Сперри и Майнера (1955) продемонстрировали, что горизонтально идущие корковые связи в большинстве случаев коротки и потому не могут обеспечить взаимодействие участков коры, находящихся на значительном отдалении друг от друга. Указанные экспериментаторы рассекали у кошек зрительную кору или вставляли в нее вертикально полоски слюды, что приводило к разделению зрительной коры на множество мелких островков, связанных между собой только через подкорковое белое вещество. Тем не менее, после этого экспериментальные животные, оправившись от операции, сохраняли способность видеть и осуществляли освоенные ими ранее зрительные дифференцировки.
Луке (Louchs R.B., 1949), Доти с соавт. (Doty R.W. et al., 1956) и др. исследователи показали, что связи между отдельными зонами зрительной коры и остальным мозгом обычно имеют форму петель, погружающихся в мозг на разную глубину. При этом отдельные колонки корковых нервных клеток могут реагировать на зрительные импульсы, возникающие при воздействии на сетчатку глаз различных по длине, интенсивности и направленности электромагнитных волн. Аналогичным образом объединяются в колонки корковые зрительные нейроны, связанные с определенными участками сетчатки правого или левого глаза, а также с периферическим глазодвигательным аппаратом. В настоящее время доказано и существование колонок глазодоминантности, а также ориентационных колонок (Хьюбел Д., 1990).
Д. Хьюбел и Т.Н. Визел (Hubel D. и WieselT.N., 1977, 1982), пользуясь микроэлектродной регистрацией биоэлектрических потенциалов нейронов, расположенных на разной глубине зрительной коры, выявили наличие между ними вертикальных возбудительных связей. При этом было установлено, что находящиеся рядом нейроны зрительной коры обычно имеют близкие по характеру функциональные свойства и объединяются в вертикальные колонки, или столбы. Указанными авторами было доказано, что нервные клетки в пределах узкой вертикальной колонки отвечают на раздражитель лишь определенной модальности, тогда как клетки соседних колонок могут автономно реагировать на зрительные сигналы той же или иной модальности. Это привело к суждению о том, что в зрительной системе группы нейронов коры реагируют главным образом на информацию, которая поступает к ним не по горизонтальным, а по вертикальным ассоциативным связям. В настоящее время доказано и существование колонок глазодоминантности, а также ориентационных колонок (Хьюбел Д., 1990).
Таким образом, зрительные импульсы, идущие от сетчатки, прежде всего, поступают в первичную зрительную кору, которая имеет не только горизонтальные, но и особенно важные вертикальные петлеобразные связи, как с вторичной зрительной корой, так и с подкорковыми центрами, влияющими на зрительные функции.
Д. Хьюбел (1990) выделял в первичной зрительной коре участки величиной приблизительно 2х2 мм. Такой участок коры, названный им модулем, обычно представляет собой группу из 500-1000 функциональных единиц. Каждый модуль имеет все механизмы, необходимые для полной обработки информации поступающей от корреспондирующего с ним участка сетчатки.
Оказалось, что если сопряженный с модулем участок сетчатки находился в области центральной ямки пятна, то такой ее участок, приходящийся на один корковый модуль (фрагмент первичной зрительной коры), как правило очень мал. Если же изучаемый фрагмент коры (модуль, имеющий аналогичную площадь), сопряжен с периферией сетчатки, то он воспринимает сигналы с участком сетчатки в несколько раз большего по площади. При этом было установлено, что модуль, на который проецируются структуры центральной ямки пятна сетчатки и модуль, сопряженный с ее периферией, нельзя признать однородными по своей структуре. Помимо разной остроты зрения по мере отдаления модуля от проекции центральной ямки сетчатки изменяются и другие параметры зрительного восприятия. Понижается, в частности, цветное и бинокулярное зрение. Вместе с тем периферические отделы сетчатки, в которых особенно значительно преобладание палочек над колбочками и сопряженные с ним корковые модули, более чувствительны к слабым источникам света и к находящимся в поле зрения перемещающимся предметам.
В процессе углубленного изучения коры больших полушарий было выявлено несколько уровней интеграции в ней зрительной информации. При этом признается особая значимость в осуществлении этого процесса ассоциативных корковых зон, в которых происходит синтез информации, добытой посредством не только зрения, но и других органов чувств. К тому же многочисленные связи коры со структурами лимбической системы во многом определяют влияние информации, поступающей с периферии через посредство анализаторов различных видов модальности, в частности, через зрительный анализатор, на состояние эмоциональной и вегетативной сфер.
- Если поле 17 (по Бродманну) обеспечивает возникновение элементарных ощущений (светоощущение, цветоощущение, элементы составляющие предмет, находящийся в обозреваемом пространстве),
- то от поля 18 зависят анализ и синтез этих ощущений и возможность расширения их количества и определенности путем рефлекторной стимуляции степени аккомодации хрусталиков и конвергенции глаз, а также изменения положения взора.
- При этом признается, что поле 19 вторичной зрительной коры уже создает возможности оптико-гностического, предметного и пространственного восприятия, способствует формированию представлений.
В процессе последующей интерпретации, возникающих в ассоциативных зонах коры уже более сложных зрительных представлений, происходит их сопоставление с имевшимися в прошлом и хранящимися в памяти зрительными впечатлениями, а также с информацией, параллельно получаемой с помощью других сенсорных систем.
Раздражение первичной зрительной коры (поле 17) обычно сопровождается появлением элементарных зрительных ощущений — фотопсий. Деструкция ее ведет к изменениям полей зрения (к вариантам неполной или полной гемианопсии, патологических скотом в гомолатеральных половинах полей зрения) и к снижению зрения, вплоть до слепоты.
Раздражение корковых полей 18 и 19, а также прилежащих к ним ассоциативных зон коры височной и теменной долей, сопровождается более сложными зрительными ощущениями по типу иллюзий или галлюцинаций, а разрушение их может обусловить нарушение ориентировки в пространстве и расстройство узнавания видимых предметов — зрительную агнозию.
Проявления зрительной агнозии особенно значительны при двустороннем нарушении функций 18, 19 корковых полей и прилежащих к ним ассоциативных корковых территорий, а также при поражении межполушарных комиссуральных связей. Больной в таких случаях может видеть и обходить препятствия, однако не узнает видимое, не имеет возможности адекватно оценить и понять окружающую обстановку. Разобраться в ней он пытается за счет сохраняющихся слуха, тактильной, температурной и других видов чувствительности.
Различают несколько видов зрительной агнозии
- агнозия на вещи — предметная,
- агнозия букв — особая форма алексии («чистая» алексия, или алексия без афазии),
- агнозия на цвета, лица и пр.
Поражение правой затылочно-теменной области часто ведет к развитию зрительной дезориентации в пространстве, к полному игнорированию его левой половины.
В связи со сложностью строения коры больших полушарий и происходящих в ней физиологических процессов изучение ее функций сопряжено с особыми трудностями. Посвятивший свою жизнь преодолению этих трудностей, американский нейрофизиолог Д. Хьюбел в 1990 г. признавал, что, несмотря на некоторые успехи в изучении преобразований зрительной информации в первичной зрительной коре, «почти ничего не известно о том, что происходит на дальнейших этапах ее переработки».
Нейропсихологами при изучении освоения человеком зрительной информации был подтвержден соматотопический порядок распространения зрительных сигналов от сетчатки до первичной зрительной коры. При этом установлено, что частичное поражение зрительных путей или первичной зрительной коры приводит к выпадению строго определенных фрагментов полей зрения, что в некоторых случаях может иметь существенное значение в топической диагностике заболеваний и травматических поражений головного мозга.
Установлено также, что раздражение первичной зрительной коры сопровождается развитием фотопсий в виде мельканий светящихся точек или пятен в определенных участках полей зрения. При поражении участков вторичной зрительной коры или прилежащих к ним ее ассоциативных зон возникают расстройства восприятия зрительных образов, в частности микро- или макрофотопсия, возможны сложные зрительные галлюцинозы или галлюцинации, а также зрительные агнозии, которые проявляются расстройством узнавания отдельных зрительных образов, нарушением способности к синтезу и обобщению получаемой зрительной информации, что ведет к дезориентации в пространстве к неадекватности представлений и поведенческих реакций.
Установлено также, что поражения отдельных участков первичной зрительной коры одного полушария не имеет сколько-нибудь серьезного значения для осуществления высших психических процессов и приводит лишь к частичным нарушениям полей зрения, в частности к возникновению патологических скотом. Однако при нарушениях функций ассоциативных зон коры теменно-затылочной области или глубинных отделов субдоминантного (чаще правого) полушария у больного возможно возникновение игнорирования противоположной половины пространства и собственного тела, которое обычно расценивается как анозогнозия и аутотопагнозия — варианты пространственной зрительной агнозии.
Если патологическим процессом оказались поражены зрительные пути с обеих сторон выше подкорковых зрительных центров, возникает центральная слепота на оба глаза, при которой остаются сохранными зрачковые рефлексы. В случаях, когда в связи с нарушением гемодинамики в сосудах вертебрально-базилярного бассейна, что на практике встречается относительно редко, избирательно поражается первичная зрительная кора обоих полушарий возможно сохранение трубчатого зрения с обеих сторон в связи с тем, что участок коры затылочной доли, на который проецируется пятно (желтое пятно) сетчатки, имеет дополнительное кровоснабжение за счет задних ветвей средних мозговых артерий.
Зрительные пути
Зрительные зоны V1 (правая и левая) передают визуальную информацию по двум первичным зрительным путям — дорсальному и вентральному.
- Дорсальный путь начинается в первичной зрительной коре (зрительная зона V1), проходит через зрительную зону V2, затем направляясь к дорсомедиальной зрительной зоне (DM или V6), зрительной зоне MT (иначе называемой V5) и в заднюю часть теменной доли коры. Дорcальный путь (канал «где?» или «как?») ассоциирован с движением, представлением о локализации объекта, управлением движениями глаз (саккады), использованием визуальной информации для оценки досягаемости объектов и доставания видимых предметов руками).
- Вентральный путь также начинается в зоне V1 и проходит через V2, но затем направляется через зрительную зону V4 к вентральной (нижней) части височной доли коры. Вентральный путь (канал «что?») связан с процессом распознавания формы, представлением об объекте, а также с долговременной памятью.
Разветвление потока зрительной информации на дорсальный («где?») и вентральный («что?») пути — иначе говоря, на каналы «действия» и «распознавания» — впервые описали Лесли Анджерлейдер и Мортимер Мишкин, и до сих пор эта гипотеза вызывает споры среди физиологов и учёных, изучающих феномен зрения. Возможно, она чрезмерно упрощает реальные процессы в зрительной коре. Она основана на обнаружении того, что оптические обманы зрения, такие как иллюзия Эббингауза, могут искажать правильное восприятие, но в случаях, когда субъект отвечает на визуальный стимул действием, таким как схватывание увиденного предмета, изображение воспринимается без искажений. Однако имеется публикация 2005 г., утверждающая, что обе кортикальные системы обработки зрительной информации — и «канал действия», и «канал распознавания» — в равной степени подвержены обману иллюзий.
Нейроны зрительной коры генерируют потенциал действия, когда визуальные стимулы воздействуют на их рецептивные поля. Рецептивное поле определяется как область зрительного поля, стимуляция которой приводит к генерации потенциала действия. Но иногда нейрон может лучше отвечать на какую-то определённую совокупность зрительных стимулов, представляемых в пределах его рецептивного поля. Это свойство называется избирательностью нейронов. В более древних зрительных областях избирательность нейронов невысока. Так, нейрон зрительной зоны V1 может возбуждаться в ответ на любой вертикальный стимул в своём рецептивном поле. Нейронам интегративных зрительных областей свойственна сложная избирательность. Например, нейроны зрительной интегративной области верхней височной борозды (у обезьян) или вентральной поверхности веретеновидной извилины на границе между затылочной и височной долями (у человека) могут возбуждаться лишь при стимуляции рецептивного поля изображениями лиц.
Кровоснабжение зрительной коры осуществляется, в основном, шпорной ветвью (лат. ramus calcarinus) медиальной затылочной артерии — разветвления конечной (корковой) части (лат. pars corticalis) задней мозговой артерии (лат. arteria cerebri posterior). Шпорная ветвь залегает в шпорной борозде (лат. sulcus calcarinus, fissura calcarina) коры головного мозга.
Вторичная область
Под термином вторичной коры подразумевают совокупность всех ассоциативных областей, которые анализируют, расшифровывают и дополняют полученную визуальную информацию с сенсорной зоны V1. Кроме того, более дальние зоны принимают непосредственное участие в формировании реакции на получаемые данные.
Обработка и анализ информации в этих зонах , возможен благодаря их строению, которое отличается от V1 наличием большого числа нейронов, которые способны выполнять сложные операции. Стоит отметить, что чем дальше зона от V1, тем большее количество сложных нейронов она содержит.
Современное представление об этой области содержит посыл о наличии в ней уровней интеграции информации. Следуя данной гипотезе, каждая зона увеличивает и уточняет данные проходящие через нее, и чем большее количество их приняло участие в обработке, тем более сложным и конечными, являются образы.
Кроме того последние исследования доказали, что некоторое число аксонов нейронов зоны V2 соединяются с центрами управления взора. Это указывает на то, что в процессе анализа и расшифровки данных вторичная зрительная кора имеет возможность дополнять ее. Для этого она рефлекторно управляет аккомодацией хрусталиков и направлением взора. Также кроме рефлекторного управления, более дальние ассоциативные зоны могут целенаправленно регулировать положение взгляда и фокусировать внимание на объекте визуализации.
Наряду с огромными усилиями, затраченными на изучение ассоциативных зон, один из самых известных нейрофизиологов считал Д.Хьюбел, что вся наука не продвинулась и на пару процентов в сторону изучении процессов анализа и обработки во вторичной структуре коры.