Введение
Проводящие пути нервной системы и состоящие из них сложные рефлекторные дуги — наиболее важный и сложный раздел неврологии. Важен он потому, что утверждает клеточную природу нервной системы (нейронная доктрина) и показывает упорядоченный характер расположения и связей нейронов (в виде рефлекторных дуг), лежащий в основе ее регулирующей функции.
При этом имеется существенное отличие от метода описательной анатомии. Последняя позволяет продемонстрировать форму, размеры и локализацию того или иного образования нервной системы, а также его принадлежность к серому или белому веществу, но совершенно не раскрывает структурную организацию нервной системы и механизмы ее функционирования.
Этот опасный для мировоззрения «отрыв» структуры от функции ликвидирует системный подход к нервной системе в виде изучения рефлекторных дуг. Здесь акцент делается именно на наличие связей нейронов, на их взаимодействие, приводящее к функционированию как самой нервной системы, так и целостного организма. Однако при этом возрастает количество мыслительных операций у обучающихся (к анализу добавляется синтез), что увеличивает трудоемкость освоения материала и его субъективную сложность. Тем не менее только изучение нервной системы как совокупности рефлекторных дуг позволяет понять ее организацию и функциональное значение. Наконец, только знание нейронных связей и взаимодействий позволяет проводить топическую диагностику поражения нервной системы, т.е. осмысленно подходить к диагностике и лечению нервных и многих других болезней и повреждений.
Как соотносятся между собой понятия «проводящий путь» и «рефлекторная дуга»? Здесь следует четко понимать, что любой проводящий путь является частью той или иной рефлекторной дуги. Поскольку в рефлекторной дуге присутствуют два главнейших звена: афферентное и эфферентное, то и проводящие пути классифицируют на афферентные и эфферентные. Учитывая иерархический принцип построения центральной нервной системы (наличие высших и подчиненных им низших нервных центров) и возможность замыкания рефлекторных дуг на уровне высших нервных центров, ясно, что и афферентные, и эфферентные проводящие пути должны быть локализованы как в периферической, так и в центральной частях нервной системы. Поскольку замыкание соматических рефлекторных дуг (соединение афферентного и эфферентного звеньев посредством вставочных нейронов) всегда происходит в центральной нервной системе, то в последних выделяют также ассоциативное звено и соответствующие ему ассоциативные проводящие пути, локализованные только в пределах центральной нервной системы.
Афферентные нервные пути проводят импульсы от рецептора до нервного центра и являются чувствительными. Афферентные нервные пути, заканчивающиеся в проекционных центрах коры полушарий большого мозга, относят к путям сознательной чувствительности. Те же афферентные пути, которые заканчиваются в подкорковых чувствительных нервных центрах, относят к путям бессознательной чувствительности.
Эфферентные нервные пути проводят импульсы от нервных центров к рабочему органу. Поскольку здесь речь идет только о соматической нервной системе, рабочим органом является скелетная мышца, поэтому эфферентные нервные пути называют двигательными. В зависимости от того, с какими нервными центрами связаны эфферентные пути, последние отвечают за выполнение как сознательных, так и бессознательных движений.
Любой проводящий путь (афферентный, ассоциативный или эфферентный) в зависимости от уровня замыкания и сложности рефлекторной дуги может быть однонейронным или многонейронным (несколько последовательно соединенных в цепь нейронов). Если рассматривать многонейронный проводящий путь как цепь, то в его пределах можно выделить звенья, представленные соответствующими нейронами. Компактно расположенные тела нейронов образуют нервные центры (узлового, ядерного или экранного типа), а аксоны, собранные в пучки, — нервные тракты. Таким образом, многонейронный проводящий путь состоит из нервных центров и трактов. В этом случае нервные центры и тракты одного и того же проводящего пути локализованы в определенных, но разных отделах нервной системы. Каждый тракт в пределах ЦНС проводит нервные импульсы обычно в одном направлении и в большинстве случаев — одного функционального содержания. Следует четко понимать, чем отличаются тракты в пределах ЦНС от пучков волокон, образующих черепные или спинномозговые нервы. Нервы содержат и афферентные, и эфферентные волокна, причем разные афферентные волокна могут проводить разные сенсорные импульсы.
В дальнейшем будет представлен материал, касающийся преимущественно соматической части нервной системы.
ОСНОВНОЙ ПРОВОДЯЩИЙ ПУТЬ ПРОИЗВОЛЬНЫХ ДВИЖЕНИЙ
4.3.1. Кора двигательной зоны
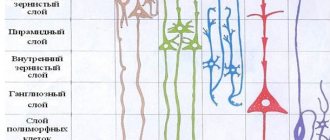
Основной проводящий путь, обеспечивающий произвольные движения, это путь, по которому нервные импульсы от двигательной зоны коры больших полушарий проходят к поперечнополосатым мышцам. Двигательной зоной коры головного мозга является, главным образом, кора предцентральной извилины (поле 4 и 6, по Бродману) и прилежащие к этой зоне ее территории. Кора здесь состоит из 6 клеточных слоев. В V слое расположены большие пирамидные клетки, описанные в 1874 г. В.А. Бецом (1834—1894). Они признаются телами центральных (верхних) двигательных нейронов, аксоны которых имеют моносинаптические связи с периферическими (нижними) мотонейронами.
4.3.2. Корково-ядерные и корково-спинномозговые связи
Нервные импульсы от центральных мотонейронов (поля 4 и 6), двигательных клеток, расположенных в прилежащих зонах лобной (поле  и теменной долей (поля 5 и 7), а также в поясной извилине (поля 23с и 24с), движутся в центробежном направлении по их аксонам, принимающим участие в формировании лучистого венца, а затем колена и передних двух третей задней ножки внутренней капсулы (рис. 4.7).
и теменной долей (поля 5 и 7), а также в поясной извилине (поля 23с и 24с), движутся в центробежном направлении по их аксонам, принимающим участие в формировании лучистого венца, а затем колена и передних двух третей задней ножки внутренней капсулы (рис. 4.7).
Колено внутренней капсулы составляют аксоны клеток центральных мотонейронов, несущие нервные импульсы к двигательным ядрам черепных нервов, расположенным в покрышке ствола мозга. Эта часть аксонов центральных двигательных нейронов формирует корково-ядерный (кортико-нуклеарный) проводящий путь. Волокна его направляются к двигательным ядрам черепных нервов, состоящим из тел периферических двигательных нейронов. Подойдя к этим ядрам, часть волокон корково-ядерного пути переходит на противоположную сторону, т.е. образуется частичный надъядерный перекрест корково-ядерных волокон. Исключение из этого правила составляют лишь волокна, идущие к нижней части ядра лицевого и к ядру подъязычного черепных нервов, так как они над уровнем соответствующих ядер совершают почти полный перекрест. В результате нижняя часть ядра лицевого нерва и ядро подъязычного черепных нервов получают нервные импульсы, как принято считать, только из противоположного полушария мозга, тогда как к остальным периферическим двигательным нейронам, расположенным в ядрах черепных нервов, нервные импульсы поступают как от противоположного большого полушария (по перекрещенным путям), так и из гомонимного полушария мозга (по неперекрещенным путям).
Аксоны центральных двигательных нейронов, составляющие передние 2/3 задней ножки внутренней капсулы, проходят в составе основания ствола мозга, и по пути следования большая их часть (приблизительно 97—98%) заканчивается у нервных клеток ретикулярной формации ствола мозга и в ядерных образованиях, относящихся к экстрапирамидной системе.
Подходя к этим образованиям, аксоны корковых клеток ветвятся, в результате большинство идущих от клеток коры эфферентных нервных импульсов возбуждает не одну, а несколько встречающихся на пути промежуточных клеток. Происходит процесс дивергенции эфферентных двигательных импульсов и, в итоге, после вовлечения в состав двигательного пути промежуточных нейронов, количество волокон, по которым продолжают перемещаться двигательные импульсы, весьма существенно увеличивается. Эфферентные двигательные пути, прошедшие и не прошедшие через промежуточные синоптические аппараты (это аксоны клеток Беца, число которых с каждой стороны — 25—30 тыс), в дальнейшем составляют пирамиды, расположенные на вентральной поверхности продолговатого мозга (подсчитано, что каждую из пирамид составляет приблизительно 1 млн нервных волокон). Пройдя пирамиды, эти волокна, названные в связи с этим в 1885 г. П, Флексигом пирамидными, на границе продолговатого и спинного мозга подвергаются частичному перекресту (decussatio pyramidum). Перешедшие на противоположную сторону пирамидные волокна (их приблизительно 80%) попадают в боковой канатик спинного мозга (funiculus lateralis) и образуют в нем боковой пирамидный путь, а двигательные волокна, оставшиеся на той же стороне (20%), входят в состав переднего канатика (funiculus anterior) и составляют в нем передний пирамидный путь. Нервные волокна, составляющие пирамидные пути, заканчиваются, подойдя к периферическим двигательным нейронам, расположенным в передних рогах сегментов спинного мозга. Все эти волокна участвуют в формировании корково-спннномозговых (кортико-спинальных) проводящих путей. Среди формирующих их нервных волокон только 2—2,5% составляют аксоны клеток Беца. Они моносинаптические, имеют наиболее толстую миелиновую оболоч-
ку и обеспечивают наибольшую скорость перемещения нервных импульсов по корково-спинальным проводящим путям.
Таким образом, пирамидный путь в настоящее время рассматривается как гетерогенная система, состоящая из нескольких подсистем с различными типами волокон, начинающихся от разных отделов мозговой коры. Подавляющее их большинство прерывается в определенных структурах подкорковых образований и ствола мозга. В связи с этим определение кортикоспинальной системы как моносинаптическойу «пирамидной» не соответствует действительности, но оно традиционное, принятое в классической неврологической литературе, поэтому в дальнейшем мы не будем избегать термина «пирамидный путь», но понимание его должно соответствовать современным представлениям.
В спинном мозге пирамидные пути спускаются вниз, при этом на уровне каждого его сегмента часть составляющих их нервных волокон заканчивается у клеток передних рогов, являющихся периферическими (нижними) двигательными нейронами.
Волокна латерального проводящего пути подходят к каждому периферическому мотонейрону, в то время как волокна переднего пирамидного пути заканчиваются только у периферических мотонейронов, которые обеспечивают на той же стороне иннервацию мышц шеи и туловища и, в частности, дыхательной мускулатуры, способствуя сохранению дыхания в случае развития у больного гемиплегии. В связи с этим периферические мононейроны, имеющие отношение к иннервации мышц шеи и туловища, получают нервные импульсы с обеих сторон, что создает условия для поддержания жизненно важных функций у больных с центральным гамипарезом.
Обобщая изложенное, можно отметить, что в передних рогах сегментов спинного мозга, где располагаются тела периферических двигательных нейронов, заканчиваются аксоны нейронов, идущие от нервных клеток, расположенных в многочисленных супраспинальных двигательных центрах. Здесь же заканчиваются и афферентные структуры, приносящие импульсы от рецепторов различных видов чувствительности, расположенных в пределах соответствующего сегмента тела.
Эфферентные и афферентные образования непосредственно или опосредованно (через вставочные нейроны) подходят к периферическим двигательным нейронам, в которых происходит суммация приходящих к ним возбуждающих и тормозных постсинаптических потенциалов (ВПСП и ТПСП). В результате синтеза импульсации, поступающей в периферический нейрон, формируется биоэлектрический заряд. Он и определяет характер образующегося в этом нейроне нервного импульса, направляющегося к сопряженным с ним мышечным волокнам.
В осуществлении и регуляции произвольных двигательных актов участвует большое количество нейротрансмиттерных систем. Уточнение характера медиаторов в разных синаптических зонах системы активных движений находится в стадии изучения. Однако уже сейчас известно, что ключевая возбуждающая нейротрансмиссия в центральных мотонейронах осуществляется в основном глутаматом и аспартатом, тогда как ГАМК и таурин способствуют снижению активности двигательных реакций. Нейротрансмиттером периферического двигательного нейрона, обеспечивающим передачу нервных импульсов в нервно-мышечных синапсах, является ацетилхолин.
Так называемый пирамидный путь не является единой, гомогенной системой волокон, он состоит из нескольких подсистем с различными связями и функциями. Прямые корковоядерные и корковоспинномозговые проекции, т.е. волокна, обеспечивающие моносинаитические связи пирамидных клеток коры больших полушарий с периферическими мотонейронами, главным образом с теми из них, которые обеспечивают иннервацию дистальных отделов конечностей, представляют собой наиболее молодую в филогенетическом отношении систему, существующую только у приматов. В настоящее время известно, что в составе проводящего пути, обычно именуемого боковым кор-тико-спинальным, или пирамидным, аксоны гигантских пирамидных клеток Беца, расположенных в коре передней центральной извилины, и имеющих мо-носинаптические связи с периферическими мотонейронами составляют всего лишь 2—2,5%.
Рис. 4.7. Пирамидная система (схема). а — пирамидный путь: 1 — кора большого мозга; 2 — внутренняя капсула; 3 — ножка мозга; 4 — мост; 5 — перекрест пирамид; 6 — латеральный корково-спинномозговой (пирамидный) путь, 7 — спинной мозг; 8 — передний корково-спинномозговой путь; 9 — чувствительные двигательные волокна периферического нерва; III, VI, VII, IX, X, XI, XII — соответствующие черепные нервы, б — конвекситальная поверхность большого полушария: двигательная зона коры (поля 4 и 6); топографическая проекция частей противоположной половины тела: 1 — ноги; 2 — туловища; 3 — руки; 4 — кисти; 5 — лица; в — горизонтальный срез через внутреннюю капсулу, расположение в ней основных проводящих путей. 1 — зрительная и слуховая лучистость; 2 — височно-мостовые и тсменно-затылочно-мостовые волокна; 3 — чувствительные таламокортикальные волокна; 4 — пирамидные пути, проводящие импульсы к мотонейронам пояснично-крес-тцового утолщения спинного мозга; 5 — пирамидные волокна, проводящие импульсы к мотонейронам грудного отдела спинного мозга; 6 — пирамидные пути, проводящие импульсы к мотонейронам шейного утолщения; 7 — корково-ядерные пути; 8 — лобно-мостовой путь; 9 — корково-таламический путь; 10 — передняя ножка внутренней капсулы; 11 — колено внутренней капсулы; 12 — задняя ножка внутренней капсулы; г — передняя поверхность ствола мозга: 1 — перекрест пирамидных путей.
4.3.3. Периферические (нижние) двигательные нейроны
В 1904 г. Шеррингтон ввел термин «конечный двигательный путь». В него он включил периферические мотонейроны и их аксоны, заканчивающиеся ветвлением, доставляющим суммарный нервный импульс к мышечным волокнам посредством нервно-мышечных синапсов, медиатором которых является аце-тилхолин-Н.
Клетки передних рогов спинного мозга представлены двумя вариантами мотонейронов: I) мотонейроны, аксоны которых имеют толстую миелиновую оболочку (А-альфа волокна), и 2) мотонейроны, аксоны которых имеют тонкую миелиновую оболочку (А-гамма волокна). Нервные клетки по аналогии с обозначением нервных волокон называются альфа- и гамма-мотонейронами. Аксоны гамма-мотонейронов направляются в мышцы, где проникают в рецеп-торные аппараты — мышечные веретена, содержащие интрафузальные (находящиеся внутри веретена) волоконца. В случаях растяжения мышцы, например при вызывании сухожильного рефлекса, происходит изменение длины и инт-рафузальных мышечных волокон: меняется конфигурация мышечных веретен, а вместе с этим происходит раздражение находящихся на них спиралевидных рецепторных структур. Возникающие в этих рецепторных структурах нервные импульсы направляются по афферентным волокнам периферических нервных структур к псевдоуниполярным клеткам спинальных ганглиев. Далее по аксонам этих клеток импульсы попадают в спинной мозг и непосредственно через коллатерали или через вставочные нейроны того же сегмента спинного мозга обеспечивают возбуждение крупных двигательных клеток (альфа-мотонейронов) передних рогов. Нервные импульсы по имеющим толстую миелиновую оболочку аксонам альфа-мотонейронов направляются к экстрафузальным волокнам, составляющим основную массу ранее подвергшихся растяжению поперечнополосатых мышц. Возникающее при этом возбуждение этих мышечных волокон вызывает напряжение или сокращение мышц, что обеспечивает пребывание в фиксированной позе или обусловливает определенный двигательный акт.
Часть дендритов нервных клеток спинальных ганглиев берет начало в мышечных сухожилиях, точнее, в расположенных в этих сухожилиях реиепторных аппаратах — сухожильных органах Гольджи. Эти рецепторы имеют высокий порог возбуждения, которое наступает лишь при чрезмерном растяжении мышечных сухожилий, угрожающем травматическим повреждением (разрывом, отрывом). В таких случаях из возбужденных сухожильных органов Гольджи в спинной мозг поступают импульсы, обеспечивающие уменьшение активности альфа-мотонейронов и снижение напряжения (тонуса) соответствующих мышц.
Кроме того, снижению мышечного тонуса при перевозбуждении альфа-мотонейронов способствуют импульсы, которые через посредство коллатералей, отходящих от их аксонов, достигают находящихся в составе переднего рога спинного мозга клеток Реншоу. Подвергшись возбуждению, эти клетки посылают тормозные биопотенциалы к альфа-мотонейронам, снижая при этом степень их напряжения.
Таким образом, периферический двигательный нейрон, наряду с влияниями, передающимися на него по эфферентным путям от корковых и экстрапирамидных структур головного мозга, испытывает воздействие импульсов, возникающих в рецепторных аппаратах на уровне того же спинального сегмента или в одноименном с этим сегментом миотоме. Все это в значительной степени способствует регуляции тонуса мышц, его оптимизации в соответствии с меняющимися требованиями, зависящими от характера выполняемых двигательных актов.
Итак, корково-мышечный путь включает центральные и периферические двигательные нейроны. К каждому периферическому нейрону поступает им-пульсания по нескольким, подчас многим нервным волокнам, идущим из коры большого мозга, из его многочисленных экстрапирамидных структур, а также из тканей одноименного сиинальному сегменту метамера тела. Часть из них представляет собой возбуждающие, часть — тормозящие биоэлектрические потенциалы. Все они в периферическом двигательном нейроне подвергаются обработке и суммации. В результате от тела периферического двигательного нейрона к мышце направляются биоэлектрические потенциалы, являющиеся результатом синтеза свойств воздействующей на него импульсации. Периферический двигательный нейрон обеспечивает иннервацию сопряженных с ним поперечнополосатых (от 5 до 1000) мышечных волокон и происходящие в них трофические процессы.
Группа мышечных волокон, иннервируемых одним периферическим мотонейроном и его аксоном, составляет двигательную единицу, при этом нервные волокна (аксон мотонейрона), идущие к мышце, обеспечивающей тонкие движения, иннервируют 5—12 мышечных волокон, тогда как крупные мышцы, осуществляющие грубые движения, получают импульсацию по нервным волокнам, каждое из которых иннервирует от 100 до 1000 их мышечных волокон. Понятие о двигательной единице как о функциональной единице нейромотор-ного аппарата было введено английским невропатологом Ч. Шеррингтоном (Sherrington Ch., 1857—1952). Мышца среднего размера имеет несколько сотен двигательных единиц. Группа периферических мотонейронов, иннервирующих одну мышцу, составляет двигательный пул.
Поражение периферических нейронов ведет к возникновению периферического паралича, который характеризуется полной обездвиженностью соответствующей части тела. Нарушение же функции центральных двигательных нейронов обусловливает центральный паралич, при котором отсутствуют произвольные движения, тогда как рефлекторные движения и синкинезии оказываются избыточными. В случае поражения части центральных двигательных нейронов, пирамидных путей или двигательных единиц, участвующих в иннервации мышц определенной части тела, в ней возникает парез, который может быть центральным или периферическим в зависимости от того, поражением элементов каких нейронов (центральных или периферических) этот парез обусловлен.
Графическая основа для изображения проводящих путей и рефлекторных дуг
Схема проводящих путей или рефлекторных дуг требует символического изображения отделов нервной системы и цепочки нейронов, расположенных в этих отделах.
Начнем с изображения нейронов. Первой в цепочке нейронов афферентного пути является ганглиозная нервная клетка, тело которой всегда расположено в периферической части нервной системы, в большинстве случаев — в чувствительном нервном узле (ганглии). По форме ганглиозная клетка выглядит либо как биполярный нейрон, либо (и чаще) как псевдоуниполярный нейрон (рис. 1).
Рис. 1. Схематическое изображение нейронов и синаптической связи. Биполярный нейрон (а); псевдоуниполярный нейрон (б): 1 — рецепторное нервное окончание, 2 — периферический отросток (дендрит), 3 — тело нейрона, 4 — центральный отросток (аксон), 5 — окончание, контактирующее со вторым нейроном в цепочке, 6 — направление импульсации. Межнейронный контакт (в): 1 — первый нейрон, 2 — синапс, 3 — второй нейрон, 4 — направление импульсации. Мотонейрон (г): 1 — тело нейрона, 2 — центральный отросток (аксон), 3 — эффекторное нервное окончание, 4 — направление импульсации
Тела коммуникационных и двигательных нейронов расположены только в центральной нервной системе: коммуникационные — в центрах либо экранного, либо ядерного типа; двигательные — только в двигательных ядрах (см. рис. 1).
Для схем большинства проводящих путей и рефлекторных дуг используется графическое изображение отделов нервной системы (рис. 2).