Активность клеток Пуркинье в коре мозжечка
Клетки Пуркинье обеспечивают выход из коры мозжечка. На их дендритах находится множество возбуждающих синапсов от параллельных волокон гранулярных клеток ( рис. 40.10 и рис. 40.11 ). Помимо этого каждая клетка Пуркинье получает обширные возбуждающие связи от лиановидного волокна , отростки которого оплетают их дендритное дерево, образуя многочисленные синаптические контакты. Сигналы от моховидных волокон вызывают в клетках Пуркинье одиночные потенциалы действия (простые спайки), тогда как ответы этих клеток на сигналы от индивидуального лиановидного волокна представляют собой ритмические разряды (сложные спайки) ( рис. 40.12 ). Поскольку частота сложных спаек низка, они не увеличивают среднюю частоту разряда клеток Пуркинье, однако, по-видимому, изменяют их чувствительность ко входам от моховидных волокон. Такие изменения могут быть долговременными и, следовательно, должны играть роль в двигательном научении .
Клетки Пуркинье получают тормозные сигналы от корзинчатых клеток и звездчатых клеток ( рис. 40.10 и рис. 40.11 ); в результате в них генерируются тормозные постсинаптические потенциалы ( рис. 40.12 ). Аксоны клеток Пуркинье спускаются через гранулярный слой в белое вещество мозжечка ( рис. 40.11 ). Большинство этих аксонов оканчиваются в каком-либо из глубинных ядер мозжечка . Некоторые клетки Пуркинье в вестибулоцеребеллуме проецируются к латеральному вестибулярному ядру . Их разряд приводит к торможению активности нейронов глубинных ядер мозжечка и латерального вестибулярного ядра . В качестве тормозного нейромедиатора из клеток Пуркинье высвобождается GABA .
На рис. 40.13 показаны некоторые нейросети мозжечка, образованные афферентными волокнами, интернейронами и клетками Пуркинье. Моховидные волокна вызывают активацию клеток-зерен и клеток Пуркинье, а также, через посредство корзинчатых и звездчатых клеток , прямое (feedforward) торможение последних ( рис. 40.13 ,а). Лиановидные волокна создают мощное возбуждение клеток Пуркинье ( рис. 40.13 ,в). Входы от моховидных волокон регулируются тормозной обратной связью с клетками Гольджи ( рис. 40.13 ,в). Сложные взаимодействия происходят между моховидными и лиановидными волокнами ( рис. 40.13 ,д). На первый взгляд может показаться парадоксальным тормозной характер выхода из коры мозжечка. Однако нужно учесть, что все входящие в него пути дают коллатерали в глубинные ядра мозжечка. Поэтому не удивительна высокая активность нейронов этих ядер. Кора мозжечка должна модулировать эту активность. Таким образом, реальный выход от мозжечка осуществляется через проекции от глубинных ядер.
История
Впервые, как несложно догадаться, клетки Пуркинье увидел человек по фамилии Пуркинье. Или Пуркине, как любят говорить пуристы. Чех Ян Эвангелиста Пуркинье был потрясающим человеком, учёным-энциклопедистом, переписывался с Гёте, состоял в ордене иллюминатов, открыл сумеречное зрение, стал одним из родоначальников дактилоскопии и создал прототип киноаппарата, несмотря на то, что был старшим современником Пушкина. Он прожил долгую жизнь, и анатомией занимался чуть более двух десятков лет. В 1837 году он описал «ганглиозные тельца» — клетки Пуркинье.
Важно Сайт школы №53
Ян Эвангелиста Пуркинье
Самое известное изображение этих ветвистых клеток получил другой великий учёный — нобелевский лауреат 1906 года, Сантьяго Рамон-и-Кахаль. На его знаменитом рисунке, растиражированном во всех учебниках, изображены клетки Пуркинье и более глубокие гранулярные клетки в мозжечке голубя.
Клетки Пуркинье (А) и гранулярные клетки (В) мозжечка голубя. Рисунок Сантьяго Рамон-и-Кахаля
Межнейронные связи мозжечка
Нейроны мозжечка не имеют прямого выхода на спинальные мотонейроны, а действуют на них через корковостволовые моторные центры.
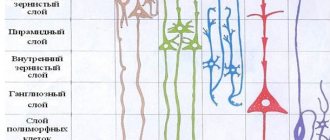
Грушевидные нейроны (клетки Пуркинье) образуют средний слой коры мозжечка и являются главной функциональной единицей, обладающей выраженной интегративной функцией. Структурной основой этой функции являются многочисленные ветвящиеся дендриты грушевидных клеток.
Клетки Пуркинье являются единственными эфферентными нейронами коры мозжечка и непосредственно связывают ее с внутримозжечковыми и вестибулярными ядрами.
Главными мозжечковыми афферентами являются лиановидные и моховидные волокна.
Лиановидные волокна. Основным источником лиановидных волокон являются нейроны нижней оливы продолговатого мозга. Информация к ним поступает от мышечных, кожных рецепторов, а также от двигательной коры больших полушарий. Каждое лиановидное волокно устанавливает синаптические контакты с дендритами (медиатор аспартат), одной клетки Пуркинье. Число синапсов может достигать трехсот, что объясняет исключительно сильное возбуждающее действие этого афферентного входа (больщой ВПСП, на вершине которого формируется 3-5 потенциалов действия).
Кроме этого, эти волокна оказывают на клетки Пуркинье и слабое тормозное влияние через корзинчатые и звездчатые клетки поверхностного слоя коры мозжечка.
По моховидным волокнам в кору мозжечка поступает информация от коры больших полушарий, проприорецепторов опорно-двигательного аппарата, вестибулярных рецепторов и ретикулярной формации.
Моховидные волокна образуют возбуждающие синапсы на дендритах клеток-зерен внутреннего слоя коры мозжечка. Через аксоны клеток-зерен этот афферентный вход оказывает непосредственное возбуждающее действие (медиатор глутамат) и опосредованное тормозное влияние на клетки Пуркинье. Активность клеток-зерен регулируется через тормозные клетки Гольджи внутреннего слоя коры мозжечка по типу возвратного торможения (медиатор ГАМК).
В мозжечке доминирует тормозной характер управления, который объясняется тем, что клетки Пуркинье, являясь тормозными нейронами, превращают возбуждающие сигналы на входе в тормозные на выходе.
Таким образом, эфферентное влияние коры мозжечка на последующее нейронное звено осуществляется по механизму сдерживания нейронной активности.
При изучении связей коры мозжечка с его ядрами выделены три симметричные вертикальные зоны.
● Клетки Пуркинье медиальной червячной зоны, а также клочка и узелка проецируются на ядра шатра.
Нейроны этих ядер связаны с моторными центрами ствола, от которых к спинальным моторным центрам идут вестибуло-, рубро- и ретикулоспинальные пути, регулирующие тонус мышц.
● Клетки Пуркинье промежуточной зоны мозжечка проецируются на шаровидное и пробковое ядра.
Аксоны нейронов этих ядер идут к красному ядру среднего мозга, от которого начинается руброспинальный путь, стимулирующий через спинальные моторные центры тонус мышц-сгибателей.
● Клетки Пуркинье латеральной зоны коры мозжечка проецируются на зубчатые ядра мозжечка, откуда пути направляются к вентролатеральному ядру таламуса, нейроны которого проецируются на моторные зоны коры большого мозга.
Итак, три парных ядра мозжечка образуют главные эфферентные выходы мозжечка на стволовые и корковые моторные центры.
Нейроны этих ядер имеют высокую фоновую активность. Они находятся под тормозным влиянием клеток Пуркинье и возбуждающим влиянием афферентных входов в мозжечок, которые по коллатеральным ответвлениям поступают в эти ядра.
Рис.1. Схема основных межнейронных связей мозжечка:
кП – клетки Пуркинье; кГ – клетки Гольджи; кз – клетки-зерна; кК – корзинчатые клетки; зк – звездчатые клетки. «+» — возбуждающие и «- « тормозные влияния.
Функции мозжечка
Функции мозжечка формируют три главных его влияния на организм: на двигательный аппарат, афферентные системы и вегетативную систему.
► Двигательные функции мозжечка заключаются в регуляции мышечного тонуса, позы и равновесия, координации позы и выполняемого движения, а также в программировании целенаправленных движений.
●Регуляция мышечного тонуса, позы и равновесия
осуществляется преимущественно древним мозжечком и частично старым мозжечком.
Получая и обрабатывая импульсацию от вестибулярных рецепторов, проприорецепторов аппарата движения, кожных, зрительных и слуховых рецепторов, мозжечок способен оценить состояние мышц, положение тела в пространстве и через ядра шатра, используя вестибуло-, ретикуло и руброспинальные пути, произвести перераспределение мышечного тонуса, изменить позу тела и сохранить равновесие.
Нарушение равновесия является наиболее характерным симптомом поражения древнего мозжечка.
●Координация позы и выполняемого целенаправленного движения
осуществляется старым и новым мозжечком. В кору этой части мозжечка поступает импульсация от рецепторов аппарата движения, а также импульсация от моторной коры. Анализируя информацию о программе движения (из моторной коры) и выполнении движения (от проприорецепторов), мозжечок способен через свои промежуточные ядра, имеющие выход на красное ядро и моторную кору, осуществить координацию позы и выполняемого целенаправленного движения, а также исправить направление движения.
Нарушение координации движения (динамическая атаксия) является наиболее характерным симптомом нарушения функции промежуточной зоны мозжечка.
●Мозжечок участвует в программировании целенаправленных движений
. Эта функция осуществляется той частью полушарий мозжечка, которая входит в латеральную зону.
Клетки Пуркинье
Кора этой части мозжечка получает импульсацию из ассоциативных зон коры больших полушарий через ядра моста. В коре нового мозжечка она перерабатывается в программу движения, которая через зубчатое ядро мозжечка и вентролатеральное ядро таламуса поступает в премоторную и моторную кору больших полушарий.
Там эта программа получает дальнейшую обработку и через пирамидную и экстрапирамидную системы реализуется как сложное целенаправленное движение.
Рис.2. Схема функций мозжечка: а — регуляция тонуса, позы и равновесия; б – координация позы и целенаправленных движений; в – программирование произвольных движений.
Контроль и коррекция более медленных программированных движений осуществляется мозжечком на основе обратной афферентации от проприорецепторов, а также от вестибулярных, зрительных и тактильных рецепторов.
Коррекция быстрых движений происходит из-за малого времени путем изменения программы в латеральном мозжечке, то сеть на основе обучения и предшествующего опыта.
► Афферентная функция мозжечка. Мозжечок имеет сложные двусторонние связи с сенсорными системами. Механизмы влияния мозжечка на сенсорные функции связаны с его влиянием на эфферентный контроль активности рецепторного аппарата и центра переключения в сенсорных системах.
В реализации влияния мозжечка на афферентные системы организма большую роль играют проекции ядер мозжечка на специфические и неспецифические ядра таламуса как главного центра переключения в сенсорных системах.
► Роль мозжечка в регуляции вегетативных функций. Мозжечок является высшим вегетативным центром. На вегетативные функции влияет старый и древний мозжечок (преимущественно червь), куда поступает часть импульсации от интерорецепторов.
Эфферентные влияния на вегетативную сферу мозжечок оказывает через ядра шатра. Они реализуются через ядра ретикулярной формации ствола и могут быть возбуждающими, тормозящими и смешанными. Конкретные механизмы переработки интероцептивной импульсации мозжечком неизвестны.
Клетки третьего типа
Гистологи выделили несколько видов клеток в терминальных отделах проводящей системы сердца. По рассматриваемой здесь классификации, клетки третьего типа буду иметь похожее строение с теми, которые составляют волокна Пуркинье в сердце. Они более объемные, по сравнению с другими водителями ритма, длинные и широкие. Толщина миофибрилл неодинакова на всех участках волокна, но сумма всех сократительных элементов получается больше, чем в обычном кардиомиоците.
Важно Аксональная полинейропатия
Теперь можно сравнить клетки третьего типа с теми, которые составляют волокна Пуркинье. Гистология (препарат, полученный из тканей на верхушке сердца) этих элементов значительно отличается. Ядро имеет практически прямоугольную форму, а сократительные волокна развиты достаточно слабо, имеют много ответвлений и соединяются между собой. Кроме того, они не ориентированы четко по длине клетки и расположены с большими промежутками. Скудное количество органелл, которые расположены вокруг миофибрилл.
Различия в частоте генерируемых импульсов и скорости их проведения, требуют филогенетически разработанного механизма синхронизации процесса сокращения во всех отделах сердца.
Мозжечок и расстройство координации движений
Моторика человека характеризуется точностью целенаправленных движений, что обеспечивается соразмерной работой многих мышечных групп, управляемых произвольно, а также автоматически. Осуществляет эту сложную многофункциональную систему многонейронный координирующий аппарат, который контролирует равновесие тела, стабилизирует центр тяжести, регулирует тонус и согласованную разнообразную деятельность мышц.
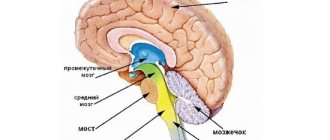
Центром координации движений является мозжечок; в функциональном отношении в нем выделяют тело мозжечка, состоящее из двух полушарий, червя и трех пар ножек.
В осуществлении произвольного движения главная роль мозжечка состоит в согласовании быстрых и медленных компонентов двигательного акта.
Это становиться возможным благодаря двусторонним связям мозжечка с мышцами и корой головного мозга. Мозжечок получает афферентные импульсы от всех рецепторов, раздражающихся во время движения (от проприорецепторов, вестибулярных, зрительных, слуховых и др.).
Получая информацию о состоянии двигательного аппарата, мозжечок оказывает влияние на красное ядро мозга, которые посылают импульсы к γ-мотонейронам спинного мозга, регулирующим мышечный тонус. Кроме того, часть афферентных импульсов через мозжечок поступает в кору больших полушарий двигательной зоны (прецентральную и лобные извилины).
Основная функция мозжечка осуществляется на подсознательном уровне.
Эфферентные импульсы от ядер мозжечка регулируют проприоцептивные рефлексы на растяжение. При мышечном сокращении происходит возбуждение проприорецептора мышц-синергистов, мышц антагонистов. В норме превращение произвольного движения в сложный рефлекс не происходит вследствие тормозного влияния мозжечковых импульсов. Поэтому при поражении мозжечка расторможенность сегментарных проприорецептивных рефлексов проявляется движениями конечностей по типу атаксии.
Мозжечок имеет многие афферентные и эфферентные связи.
Задний спинно-мозжечковый путь (путь Флексига). Первый нейрон заложен в спинномозговом ганглии, аксон в составе заднего корешка через задний рог подходит к клеткам колонки Кларка. Волокна этих вторых нейронов направляются в наружные слои задней части бокового канатика своей стороны, поднимаются вдоль всего спинного мозга и на уровне продолговатого мозга в составе нижней мозжечковой ножки входят в червь мозжечка.
Этот путь обозначается как задний спинно-мозжечковый путь. В коре червя мозжечка находится третий нейрон, который контактирует с грушевидными нейронами коры полушария мозжечка. Аксоны последних идут к зубчатому ядру. Волокна этого пятого нейрона входят в состав верхней ножки мозжечка.
Правая и левая верхние ножки мозжечка перекрещиваются — перекрест Вернике и заканчивается у клеток красного ядра противоположной стороны. Аксоны клеток красного ядрра сразу же направляются на противоположную сторону среднего мозга и образуют вентральный перекрест в покрышке среднего мозга — перекрест Фореля, проходят в составе бокового канатика спинного мозга, достигают клеток передних рогов.
Совокупность аксонов клеток красного ядра называется пучок Монакова.
Передний спинно-мозжечковый путь Говерса. Первый нейрон расположен в спинномозговом ганглии, второй нейрон — клетка заднего рога, однако аксоны ее переходят на противоположную сторону и направляются вверх по спинному мозгу, в передней части бокового канатика, проходят через продолговатый мозг, мост мозга, на уровне верхнего мозгового паруса переходят на противоположную сторону и в составе верхней ножки мозжечка достигают клеток ядер мозжечка.
Афферентные проприоцептивные импульсы мозжечок получает не только по путям Флексига и Говерса, они поступают также и по аксонам клеток ядер тонкого и клиновидного пучков, часть которых идет через нижние ножки мозжечка к его червю.
Кроме того, к мозжечку в составе нижней ножки идут аксоны клеток вестибулярных ядер — в основном от преддверного латерального ядра Дейтерса, они заканчиваются в ядре ската мозжечка.
Волокна клеток этого ядра в составе верхней и, возможно, нижней ножек мозжечка подходят к клеткам ретикулярной формации ствола мозга и преддверному латеральному ядру, от которых проводники образуют нисходящие тракты — преддверно-спинномозговой и ретикулярно-спинномозговой, заканчивающиеся у клеток передних рогов спинного мозга.
По этому пути осуществляется регуляция равновесия тела.
От мозжечка через преддверное латеральное ядро устанавливаются связи и с ядрами глазодвигательных нервов (в составе медиального продольного пучка).
Мозжечок имеет многочисленные связи почти со всеми долями мозга. Выделяют 2 массивных пучка.
Лобно-мосто-мозжечковый — совокупность аксонов клеток преимущественно передних отделов верхней и средней лобных извилин. В глубине доли они собираются в компактный пучок и образуют переднюю ножку внутренней капсулы.
Затем проходят в сновании ножки мозга и заканчиваются синапсом у клеток моста мозга. Аксоны вторых нейронов переходят на противоположную сторону моста и в составе средней ножки мозжечка входят в его полушарие, контактируя с клетками коры мозжечка. Отростки этих нейронов подходят к зубчатому ядру. Волокна клеток зубчатого ядра в составе верхней ножки мозжечка достигают красного ядра противоположной стороны и по ретикулярно-спинномозговому тракту проводят импульсы, регулирующие позы человека в вертикальном положении, в частности стояние и ходьбу.
Затылочно-височно-мосто-мозжечковый путь — первые его нейроны расположены в коре затылочной и височной доли; аксоны их собираются в подкорковом белом веществе, затем в составе заднего бедра внутренней капсулы идут в основании среднего мозга до ядер моста мозга своей стороны.
Аксоны клеток моста переходят на противоположную сторону и по средней ножке достигают коры мозжечка. Волокна этих клеток подходят к зубчатому ядру, которое имеет связи со стволом мозга. С помощью этих трактов обеспечивается координация работы мозжечка с органами зрения и слуха.
В конечном итоге, существующие перекресты мозжечковых афферентных и эфферентных систем приводят к гомолатеральной связи одного полушария мозжечка и конечностей.
При поражении полушария мозжечка расстройства его функции возникают на одноименной половине тела. Очаги в боковом канатике спинного мозга также вызывают мозжечковые расстройства на своей половине тела. Полушария головного мозга соединены с противоположными полушариями мозжечка.
Поэтому при поражении полушарий большого мозга или красного ядра мозжечковые расстройства выявляются на противоположной стороне.
Определение
Клетки Пуркинье (Purkinje cells) названы в честь чешского ученого, впервые обнаружившего и описавшего их – Яна Пуркинье (Jan Purkyně). Форма клетки напоминает грушу. От нее отходит несколько дендритов, которые в свою очередь выпускают многочисленные ветви с образованием тысяч синапсов. Клетки Пуркинье – это корковый слой мозжечка – отдела, который находится ниже затылочного участка мозга головы.
: управление двигательной координацией, поддержание равновесия. Благодаря этому небольшому участку (10% общего объема) головного мозга человек совершает все движения: играет в футбол, удерживает и подносит ко рту столовые приборы, выводит рукой письменные знаки, печатает на клавиатуре, перемещает глаза, следя за объектами внешнего мира.
Важно Успокоительные травяные чаи: что выбрать?
Оценить значимость этой мозговой структуры можно, представив с какой тщательностью должны быть синхронизированы движения, чтобы они были точными, слаженными и плавными. Мозжечок корректирует активность скелетных мышц, чтобы в результате добиться целенаправленной точности движения – до миллиметра (в пространственной плоскости), до доли секунды (во временных рамках).
Мозжечок управляет движениями головы, конечностей, тела, глаз, параллельно получая обратные сведения, отправляемые нервными окончаниями периферической . Сопоставляя желаемый и действительный результат, он корректирует моторную деятельность. Все процессы происходят мгновенно и незаметно для человека.
ПОНЯТИЕ АТАКСИИ, ВИДЫ АТАКСИЙ
Атаксия — форма беспорядочного движения, taxis — от греч. порядок, а- отрицание. Возникает при нарушении согласованности действия мышечных групп-агонистов (непосредственно осуществляющих движение), антагонистов (в какой-то фазе противодействующих агонистам), синергистов (помогающих работе то агонистов, то антагонистов).
Движения утрачивают слаженность, точность, плавность, соразмерность и часто не достигают цели. Мышечная сила у такого больного остается достаточной, у него достаточная, у него нет парезов.
Патогенетическая сущность атаксии состоит:
Нарушение реципрокной иннервации. Механизм реципрокного (сопряженного) торможения спинномозговых двигательных центров заключается в следующем: аксоны рецепторных клеток (в спинальных ганглиях) в спинном мозге делятся на ветви, одни из них возбуждают мотонейроны мышц-сгибателей, а другие — контактируют с вставочными клетками, которые оказывают тормозное влияние на клетки мышц-разгибателей.
В сложной интегративной функции этого механизма участвуют также и мозжечковые импульсы.
2. Прекращение проприоцептивной сигнализации (от мышечных веретен, сухожильных телец Гольджи) по тому или иному восходящему афферентному пути. Перестает поступать информация о степени напряжения мышц в каждый данный момент, о результатах адаптационных эффектов функциональных систем.
Расстраивается та сторона двигательной функции, которую обозначают обратной афферентацией.
Виды атаксий.
1. Сенситивная атаксия, связана с одновременным страданием координации движений и мышечно-суставного чувства. При выраженной атаксии в верхней конечности затруднено выполнение самых простых действий. В покое в пальцах кисти иногда наблюдаются непроизвольные движения, напоминающие атетоз — псевдоатетоз. Нарушена координация движений также и в нижних конечностях, что подтверждается мимопопаданием и толчкообразными движениями при выполнении пяточно-коленной пробы.
Мышечный тонус в пораженных конечностях оказывается пониженным и в мышцах-сгибателях, и в разгибателях. В положении стоя отмечается пошатывание, особенно при проведении пробы Ромберга.
Передвижение становится неуверенным, больной ходит с опущенной головой, контролируя акт ходьбы с помощью зрения. Таким образом, сенситивная атаксия всегда сочетается с расстройством глубокой чувствительности и функциональным разобщением отдельных сегментов конечностей с высшими зонами мозга. Другой характерной чертой этого вида атаксии является усиление ее при выключении контроля зрительного анализатора.
Сенситивная атаксия прии поражении задних канатиков нижней половины спинного мозга (например, при сифилисе, фуникулярном миелозе — В12) может сопровождаться исчезновением глубоких рефлексов на нижних конечностях, что объясняется дегенерацией коллатералей волокон тонкого пучка, являющихся афферентной частью дуги глубоких рефлексов.
Мозжечковая атаксия.
Связана с поражением мозжечковых систем. Учитывая, что червь мозжечка принимает участие в регуляции сокращения мышц туловища, а кора полушарий — дистальных отделов конечностей, различают две формы мозжечковой атаксии.
— Статико-локомторная — при поражении червя мозжечка расстраиваются в основном стояние и ходьба.
Больной стоит с широко расставленными ногами, покачивается. При ходьбе туловище отклоняется в стороны, походка напоминает походку пьяного. Особенно затруднены повороты. Отклонение при ходьбе наблюдаются в сторону мозжечкового поражения. Устойчивость проверяется в позе Ромберга.
При поражении мозжечковых структур больной в этой позе покачивается в соответствующую сторону; при пошатывании в переднезаднем направлении — характерно для поражения передних отделов червя мозжечка.
Исследуют ходьбу больного по прямой, а также фланговую походку — шаговые движения в сторону. При этом обращают внимание на четкость шага и на возможность быстрой остановки при внезапной команде. При поражении мозжечковых структур нарушается сочетание простых движений, составляющих последовательную цепь сложных двигательных актов. Это обозначается как асинергия, определяется с помощью пробы Бабинского.
— Динамическая атаксия — при ней нарушается выполнение различных произвольных движений конечностями.
Этот вид атаксии в основном зависит от поражения полушарий мозжечка. Это обнаруживается при исследовании движений верхних конечностей, например при выполнении пальценосовой, пяточно-коленной, пробы на диадохокинез и т.д.
Кроме нарушения этих проб с движениями конечностей, при поражении мозжечка расстраивается речь — в результате инкоординации речедвигательной мускулатуры речь теряет плавность, становится взрывчатой, ударения ставятся не на нужных слогах — скандирующая речь; изменяется почерк -почерк становится неровным, крупным — мегалография.
Наблюдается нистагм — ритмическое подергивание глазных яблок при взгляде в стороны или вверх — своего рода интенционный тремор глазодвигательных мышц. При поражении мозжечковых структур плоскость нистагма совпадает с направлением произвольных движений глазных яблок — при взгляде в стороны нистагм горизонтальный, при взгляде вверх-вниз — вертикальный.
Иногда нистагм является врожденным. Такой нистагм обычно имеется не только при отведении глазных яблок в стороны (при напряжении), но и при взгляде прямо («спонтанный нистагм»).
При поражении мозжечковых систем может изменяться мышечный тонус.
Наиболее часто наблюдается мышечная гипотония: мышцы становятся дряблыми, вялыми, возможна гипермобильность суставов. При этом могут снижаться глубокие рефлексы.
Координация движений нарушается при поражении лобной и височной долей и их проводников. В таком случае расстраивается ходьба и стояние, туловище отклоняется кзади и в сторону, противоположную очагу. Выявляется мимопопадание в руке и ноге (гемиатаксия). При таком виде нарушения координации обнаруживаются и другие признаки поражения соответствующих долей больших полушарий.
Корковая атаксия. Координация движений нарушается при поражении лобной и височной долей и их проводников. В таком случае расстраивается ходьба и стояние, туловище отклоняется кзади и в сторону, противоположную очагу. Выявляется мимопопадание в руке и ноге (гемиатаксия). При таком виде нарушения координации обнаруживаются и другие признаки поражения соответствующих долей больших полушарий.
4. Вестибулярная атаксия — возникает при нарушении функции вестибулярного анализатора, в частности его проприорецепторов в лабиринте.
При ней расстраивается равновесие тела, во время ходьбы больной отклоняется в сторону пораженного лабиринта. Характерны системное головокружение, тошнота, а также горизонтально-ротаторный нистагм. На стороне пораженного лабиринта может нарушаться слух.
Таким образом, расстройство координации произвольных движений наблюдается при поражении как самого мозжечка, так и проводников, по которым приводятся к нему импульсы от мышц, полукружных каналов внутреннего уха и коры головного мозга и отводятся от мозжечка к двигательным нейронам мозгового ствола и спинного мозга.
Больные с поражением мозжечковых систем в покое обычно никаких патологических проявлений не обнаруживают. Различные виды инкоординации появляются у них только при напряжении мышц.