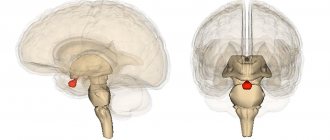
Средняя линия перед моторной корой головного мозга
Некоторые двигательные области в коре головного мозга человека. Дополнительная моторная зона показана розовым цветом. Автор изображения: Паскари
В дополнительная моторная зона
(
SMA
) является частью коры головного мозга приматов, которая способствует управлению движением. Он расположен на средней линии поверхности полушария прямо перед (перед) представлением ноги первичной моторной коры. У обезьян SMA содержит приблизительную карту тела. У людей карта тела не видна. Нейроны в SMA проецируются непосредственно на спинной мозг и могут играть роль в прямом управлении движением. Возможные функции, приписываемые SMA, включают постуральную стабилизацию тела, координацию обеих сторон тела, например, во время бимануального действия, контроль движений, которые генерируются внутренне, а не запускаются сенсорными событиями, и контроль последовательности движений. . Все эти предложенные функции остаются гипотезами. Точная роль или роли SMA пока не известны.
Об открытии SMA и ее связи с другими моторными областями коры см. Основную статью о моторная кора.
Субрегионы
По крайней мере, шесть областей теперь распознаются в более крупном регионе, когда-то определяемом как SMA. Эти подразделения наиболее подробно изучены в мозге обезьян. Самая передняя часть в настоящее время обычно называется пре-SMA.[1][2][3] Он редко связан со спинным мозгом или первичной моторной корой или вообще не связан со спинным мозгом, а также имеет обширные связи с префронтальными областями.[1][4][5][6][7]
В дополнительное глазное поле (SEF) представляет собой относительно переднюю часть SMA, которая при стимуляции вызывает движения головы и глаз и, возможно, движения конечностей и туловища.[8][9][10][11]
Дам и Стрик[5] выдвинуто на основе цитоархитектура и соединения со спинным мозгом, что часть SMA в поясная борозда, в средней части полушария, может быть разделен на три отдельные области, поясные двигательные области. Функции двигательных зон поясной извилины систематически не изучены.
Собственно SMA у обезьян теперь ограничивается областью на макушке полушария и частично распространяется на медиальную стенку, непосредственно перед представлением первичной двигательной ноги. Собственно SMA проецируется непосредственно на спинной мозг и, следовательно, является одной из основных выходных областей корковой моторной системы.[5][12][13][14][15][16]
Недавно Zhang et al.[17] исследовали функциональные подразделения медиальной SFC на основе связи всего мозга, охарактеризованной на основе большого набора данных фМРТ в состоянии покоя. Помимо воспроизведения границ между SMA и preSMA, текущие результаты подтверждают функциональную разницу между задним и передним пре-SMA. В отличие от задней пре-SMA, передняя пре-SMA связана с большей частью префронтальных, но не сомато-моторных областей. В целом, SMA прочно связана с таламусом и эпиталамус, задний пре-SMA до скорлупа, паллидум, а также STN и передняя пре-SMA к хвостатое ядро, причем хвостатые части демонстрируют значительную асимметрию полушарий.
Особенности строения коры мозга
Анатомическое строение коры больших полушарий обусловливает ее особенности и позволяет выполнять возложенные на нее функции. Кора головного мозга владеет следующим рядом отличительных черт:
- нейроны в ее толще располагаются послойно;
- нервные центры находятся в конкретном месте и отвечают за деятельность определенного участка организма;
- уровень активности коры зависит от влияния ее подкорковых структур;
- она имеет связи со всеми нижележащими структурами центральной нервной системы;
- наличие полей разных по клеточному строению, что подтверждается гистологическим исследованием, при этом каждое поле отвечает за выполнение какой-либо высшей нервно деятельности;
- присутствие специализированных ассоциативных областей позволяет устанавливать причинно-следственную связь между внешними раздражителями и ответом организма на них;
- способность к замещению поврежденных участков близлежащими структурами;
- этот отдел мозга способен сохранять следы возбуждения нейронов.
Большие полушария головного мозга состоят главным образом из длинных аксонов, а также содержит в своей толще скопления нейронов, образующих наибольшие ядра основания, которые входят в состав экстрапирамидальной системы.
Как уже говорилось, формирование коры мозга происходит еще во время внутриутробного развития, причем вначале кора состоит из нижнего слоя клеток, а уже в 6 месяцев ребенка в ней сформированы все структуры и поля. Окончательное становление нейронов происходит к 7-летнему возрасту, а рост их тел завершается в 18 лет.
Интересен тот факт, что толщина коры не равномерна на всей протяженности и включает в себя разное количество слоев: например, в области центральной извилины она достигает своего максимального размера и насчитывает все 6 слоев, а участки старой и древней коры имеют 2-х и 3-х слойное строение соответственно.
Нейроны этой части мозга запрограммированы на восстановление поврежденного участка посредством синоптических контактов, таким образом каждая из клеток активно старается восстановить поврежденные связи, что обеспечивает пластичность нейронных корковых сетей. Например, при удалении или дисфункции мозжечка, нейроны, связывающие его с конечным отделом, начинают прорастать в кору больших полушарий.
анализатора, замыкательного аппарата условно-рефлексных связей и рабочего прибора. Слабость замыкательной функции коры и следовых проявлений можно наблюдать у детей с выраженной умственной отсталостью, когда образовавшиеся условные связи между нейронами хрупки и ненадежны, что влечет за собой трудности в обучении.
Кора головного мозга включает в себя 11 областей, состоящих из 53 полей, каждому из которых в нейрофизиологии присвоен свой номер.
Области и зоны коры
Кора относительно молодая часть ЦНС, развывшаяся из конечного отдела мозга. Эволюционно становление этого органа происходило поэтапно, поэтому ее принято разделять на 4 типа:
- Архикортекс или древняя кора в связи с атрофией обоняния превратился в гиппокамповую формацию и состоит из гиппокампа и сопряженных ему структур. С помощью ее регулируется поведение, чувства и память.
- Палеокортекс или старая кора, составляет основную часть обонятельной зоны.
- Неокортекс или новая кора имеет толщину слоя около 3—4 мм. Является функциональной частью и совершает высшую нервную деятельность: обрабатывает сенсорную информацию, отдает моторные команды, а также в ней формируется осознанное мышление и речь человека.
- Мезокортекс является промежуточным вариантом первых 3 типов коры.
Кора головного мозга имеет сложную анатомическую структуру и включает в себя сенсорные клетки, моторные нейроны и интернероны, обладающих способностью останавливать сигнал и возбуждаться в зависимости от поступивших данных. Организация этой части мозга построена по колончатому принципу, в котором колонки делаться на микромодули, имеющие однородное строение.
Основу системы микромодулей составляют звездчатые клетки и их аксоны, при этом все нейроны одинаково реагируют на поступивший афферентный импульс и посылают также синхронно в ответ эфферентный сигнал.
Формирование условных рефлексов, обеспечивающих полноценное функционирование организма, и происходит благодаря связи головного мозга с нейронами, расположенными в различных частях тела, а кора обеспечивает синхронизацию умственной деятельности с моторикой органов и областью, отвечающей за анализ поступающих сигналов.
Передача сигнала в горизонтальном направлении происходит через поперечные волокна, находящиеся в толще коры, и передают импульс от одной колонки к другой. По принципу горизонтальной ориентации кору мозга можно поделить на следующие области:
- ассоциативная;
- сенсорная (чувствительная);
- моторная.
При изучении этих зон применялись различные способы воздействия на нейроны, входящие в ее состав: химическое и физическое раздражение, частичное удаление участков, а также выработка условных рефлексов и регистрация биотоков.
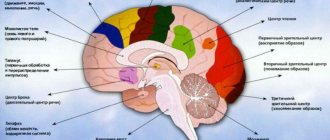
Ассоциативная зона связывает поступившую сенсорную информацию с полученными ранее знаниями. После обработки формирует сигнал и передает его в двигательную зону. Таким образом она участвует в запоминании, мышлении и обучении новым навыкам. Ассоциативные участки коры головного мозга расположены в близости с соответствующей сенсорной зоной.
Чувствительная или сенсорная зона занимает 20% коры головного мозга. Она также состоит из нескольких составляющих:
- соматосенсорной, расположенной в теменной зоне отвечает за тактильную и вегетативную чувствительность;
- зрительной;
- слуховой;
- вкусовой;
- обонятельной.
Импульсы от конечностей и органов осязания левой стороны тела, поступают по афферентным путям в противоположную долю больших полушарий для последующей обработки.
Нейроны моторной зоны возбуждаются при помощи импульсов, поступивших от клеток мускулатуры, и находятся в центральной извилине лобной доли. Механизм поступления данных схож с механизмом сенсорной зоны, так как двигательные пути образуют перехлест в продолговатом мозге и следуют в расположенную напротив моторную зону.
Кора больших полушарий образована несколькими слоями нейронов. Характерной особенностью этой части мозга является большое количество морщин или извилин, благодаря чему ее площадь во много раз превосходит площадь поверхности полушарий.
Корковые архитектонические поля определяют функциональное строение участков коры головного мозга. Все они различны по морфологическим признакам и регулируют разные функции. Таким образом выделяется 52 различных поля, расположенных на определенных участках. По Бродману это разделение выглядит следующим образом:
- Центральная борозда разделяет лобную долю от теменной области, впереди нее пролегает предцентральная извилина, а сзади — позадицентральная.
- Боковая борозда отгораживает теменную зону от затылочной. Если развести ее боковые края то внутри можно рассмотреть ямку, в центре которой имеется островок.
- Теменно-затылочная борозда отделяет теменную долю от затылочной.
В предцентральной извилине расположено ядро двигательного анализатора, при этом к мышцам нижней конечности относятся верхние части передней центральной извилины, а к мышцам полости рта, глотки и гортани – нижние.
Правосторонняя извилина образует связь с двигательным аппаратом левой половины тела, левосторонняя – с правой частью.
В позадицентральной извилине 1 доли полушария содержится ядро анализатора тактильных ощущений и она также связана с противолежащей частью тела.
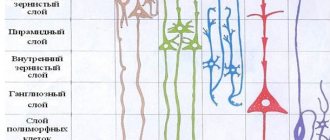
Клеточные слои
Кора головного мозга осуществляет свои функции посредством нейронов, находящихся в ее толще. Причем количество слоев этих клеток может отличаться в зависимости от участка, габариты которых также разнятся по размеру и топографии. Специалисты выделяют следующие слои коры головного мозга:
- Поверхностный молекулярный сформирован в основном из дендритов, с небольшим вкраплением нейронов, отростки которых не покидают границы слоя.
- Наружный зернистый состоит из пирамидальных и звездчатых нейронов, отростки которых связывают его со следующим слоем.
- Пирамидальный образован пирамидными нейронами, аксоны которых направлены вниз, где обрываются или образуют ассоциативные волокна, а дендриты их соединяют этот слой с предыдущим.
- Внутренний зернистый слой сформирован звездчатыми и малыми пирамидальными нейронами, дендриты которых уходят в пирамидальный слой, а также его длинные волокна уходят в верхние слои или спускаются вниз в белое вещество мозга.
- Ганглионарный состоит из крупных пирамидальных нейроцитов, их аксоны выходят за пределы коры и связывают различные структуры и отделы ЦНС между собой.
Мультиформный слой сформирован всеми видами нейронов, а их дендриты ориентированы в молекулярный слой, а аксоны пронизывают предыдущие слои или выходят за пределы коры и образуют ассоциативные волокна, образующие связь клеток серого вещества с остальными функциональными центрами головного мозга.
Функции
Penfield и Уэлч[18] в 1951 году впервые описал СМА в головном мозге обезьяны и человека как изображение тела на медиальной стенке полушария. Вулси и его коллеги[19] в 1952 г. подтвердил СМА в головном мозге обезьяны, описав ее как грубую соматотопную карту с ногами в задней части и лицом в передней части. Было обнаружено, что изображения разных частей тела сильно пересекаются. Стимуляция многих участков вызывала двусторонние движения, а иногда и движения всех четырех конечностей. Эта перекрывающаяся соматотопическая карта в SMA была подтверждена многими другими.[2][13][20][21][22]
Были предложены четыре основные гипотезы для функции SMA: контроль устойчивости позы во время стойки или ходьбы,[18] согласование временных последовательностей действий,[23][24][25][26][27][28][29][30] бимануальная координация,[31][32] и инициирование внутреннего движения, а не движения, управляемого стимулом.[3][29][30][33] Однако данные, как правило, не подтверждают исключительную роль SMA в любой из этих функций. Действительно, СМА явно активна во время непоследовательных, однообразных движений и движений по стимулам.[34]
Для произвольного движения человека роль SMA была выяснена: ее активность генерирует ранний компонент Bereitschaftspotential (БП) или потенциал готовности БП1 или БП на раннем этапе.[35] Роль SMA была дополнительно подтверждена Cunnington et al. 2003 г.,[36] показывает, что собственно SMA и пре-SMA активны до произвольного движения или действия, а также поясная двигательная зона (CMA) и передняя часть средней поясной коры (aMCC). Недавно путем интеграции одновременно полученных ЭЭГ и фМРТ было показано, что SMA и aMCC имеют сильные реципрокные связи, которые действуют, чтобы поддерживать активность друг друга, и что это взаимодействие опосредуется во время подготовки к движению в соответствии с амплитудой потенциала Bereitschafts.[37]
СМА в мозгу обезьяны может усиливать движение, особенно сложное, такое как лазание или прыжок.[38][39][40] Это предположение было основано на исследованиях, в которых стимуляция во временной шкале, имеющей отношение к поведению, вызывала сложные движения всего тела, напоминающие лазание или прыжки. Эта гипотеза согласуется с предыдущими гипотезами, включая участие SMA в стабилизации позы, во внутренне генерируемых движениях, в бимануальной координации и в планировании последовательности движений, потому что все эти функции в значительной степени задействованы в сложной локомоции. Гипотеза локомоции — это пример интерпретации моторной коры головного мозга с точки зрения основного поведенческого репертуара, из которого возникают абстрактные функции управления, — подход, подчеркнутый Грациано и коллеги.[38]
Роль коры больших полушарий
В задней центральной извилине, позади от центральной борозды, располагается зона кожной и суставно-мышечной чувствительности. Здесь воспринимаются и анализируются сигналы, возникающие при касании к нашему телу, при воздействии на него холода или тепла, болевых воздействиях.
В противоположность этой зоне — в передней центральной извилине, спереди от центральной борозды, расположена двигательная зона. В ней выявлены участки, которые обеспечивают движения нижних конечностей, мышц туловища, рук, головы. При раздражении этой зоны электротоком возникают сокращения соответствующих групп мышц. Ранения или другие повреждения коры двигательной зоны влекут за собой паралич мышц тела.
В височной доле находится слуховая зона. Сюда поступают и здесь анализируются импульсы, возникающие в рецепторах улитки внутреннего уха. Раздражения участков слуховой зоны вызывают ощущения звуков, а при поражении их болезнью утрачивается слух.
Зрительная зона расположена в коре затылочных долей полушарий. При ее раздражении электрическим током во время операций на мозге человек испытывает ощущения вспышек света и темноты. При поражении ее какой-либо болезнью ухудшается и теряется зрение.
Вблизи боковой борозды расположена вкусовая зона, где анализируются и формируются ощущения вкуса на основании сигналов, возникающих в рецепторах языка. Обонятельная зона расположена в так называемом обонятельном мозге, у основания полушарий. При раздражении этих зон во время хирургических операций или при воспалении люди ощущают запах или вкус каких-либо веществ.
Чисто речевой зоны не существует. Она представлена в коре височной доли, нижней лобной извилине слева, участках теменной доли. Их поражения болезнями сопровождаются расстройствами речи.
Большие полушария головного мозга занимают около 80% объема черепной коробки, и состоят из белого вещества, основа которого состоит из длинных миелиновых аксонов нейронов. Снаружи полушария покрывает серое вещество или кора головного мозга, состоящая из нейронов, безмиелиновых волокон и глиальных клеток, которые также содержатся в толще отделов этого органа.
Поверхность полушарий условно делится на несколько зон, функциональность которых заключается в управлении организмом на уровне рефлексов и инстинктов. Также в ней находятся центры высшей психической деятельности человека, обеспечивающие сознание, усвоение поступившей информации, позволяющей адаптироваться в окружающей среде, и через нее, на уровне подсознания, посредством гипоталамуса контролируется вегетативная нервная система (ВНС), управляющая органами кровообращения, дыхания, пищеварения, выделения, размножения, а также метаболизмом.
Для того чтобы разобраться что такое кора мозга и каким образом осуществляется ее работа, требуется изучить строение на клеточном уровне.
Рекомендации
- ^ аб
Он, С.К., Дум, Р.П. и Стрик, П.Л. (1995). «Топографическая организация кортикоспинальных проекций лобной доли: двигательные зоны на медиальной поверхности полушария».
J. Neurosci
.
15
(5): 3284–3306. Дои:10.1523 / JNEUROSCI.15-05-03284.1995.CS1 maint: несколько имен: список авторов (ссылка на сайт) - ^ аб
Луппино, Дж., Мателли, М., Камарда, Р. М., Галлезе, В. и Риццолатти, Г. (1991). «Множественные репрезентации движений тела в мезиальной области 6 и прилегающей поясной коре головного мозга: исследование внутрикортикальной микростимуляции на макаках».
J. Comp. Neurol
.
311
(4): 463–482. Дои:10.1002 / cne.903110403. PMID 1757598.CS1 maint: несколько имен: список авторов (ссылка на сайт) - ^ аб
Мацудзака Ю., Айзава Х. и Танджи Дж. (1992). «Двигательная область растет в дополнительную двигательную область (предварительная двигательная область) у обезьяны: нейронная активность во время выученной двигательной задачи».
J. Neurophysiol
.
68
(3): 653–662. Дои:10.1152 / ян.1992.68.3.653. PMID 1432040.CS1 maint: несколько имен: список авторов (ссылка на сайт) - Бейтс, Дж. Ф. и Гольдман-Ракич, П. С. (1993). «Префронтальные связи медиальных моторных областей у макаки-резуса». J. Comp. Neurol
.
336
(2): 211–228. Дои:10.1002 / cne.903360205. PMID 7503997. - ^ абc
Дум Р. П. и Стрик П. Л. (1991). «Происхождение кортикоспинальных проекций из премоторных областей лобной доли».
J. Neurosci
.
11
(3): 667–689. Дои:10.1523 / JNEUROSCI.11-03-00667.1991. - Лу, М.Т., Престон, Дж. Б. и Стрик, П. Л. (1994). «Взаимосвязи между префронтальной корой и премоторными областями лобной доли». J. Comp. Neurol
.
341
(3): 375–392. Дои:10.1002 / cne.903410308.CS1 maint: несколько имен: список авторов (ссылка на сайт) - Луппино, Г., Мателли, М., Камарда, Р. и Риццолатти, Г. (1993). «Кортикокортикальные связи области F3 (SMA-собственно) и области F6 (pre-SMA) у макак». J. Comp. Neurol
.
338
(1): 114–140. Дои:10.1002 / cne.903380109. PMID 7507940.CS1 maint: несколько имен: список авторов (ссылка на сайт) - Чен Л.Л. и Уолтон М.М. (2005). «Движение головы, вызванное электростимуляцией в дополнительном поле глаза макаки-резуса». J. Neurophysiol
.
94
(6): 4502–4519. Дои:10.1152 / ян.00510.2005. - Руссо, Г.С. и Брюс, К.Дж. (2000). «Дополнительное поле глаза: представление саккад и взаимосвязь между полями нервных реакций и вызванными движениями глаз». J. Neurophysiol
.
84
(5): 2605–2621. Дои:10.1152 / jn.2000.84.5.2605. - Шлаг, Дж. И Шлаг-Рей, М. (1987). «Доказательства дополнительного глазного поля». J. Neurophysiol
.
57
: 179–200. Дои:10.1152 / jn.1987.57.1.179. - Теховник, Э. И Ли, К. (1993). «Дорсомедиальная лобная кора головного мозга макаки-резуса: топографическое изображение саккад, вызванных электрической стимуляцией». Exp. Мозг Res
.
96
(3): 430–442. Дои:10.1007 / bf00234111. - Галеа, М. И Дариан-Смит, я (1994). «Множественные популяции кортикоспинальных нейронов у макак определяются их уникальным корковым происхождением, спинномозговыми окончаниями и связями». Цереб. Кора
.
4
(2): 166–194. Дои:10.1093 / cercor / 4.2.166. - ^ аб
Макферсон, Дж., Марангоз, К., Майлз, Т.С. и Wiesendanger, M (1982). «Микростимуляция дополнительной двигательной области (SMA) у бодрствующей обезьяны».
Exp. Мозг Res
.
45
(3): 410–416. Дои:10.1007 / bf01208601.CS1 maint: несколько имен: список авторов (ссылка на сайт) - Мюррей, Э.А. И Коултер, Дж. Д. (1981). «Организация кортикоспинальных нейронов у обезьяны». J. Comp. Neurol
.
195
(2): 339–365. Дои:10.1002 / cne.901950212. - Нудо, Р.Дж. И Мастертон, Р. Б. (1990). «Нисходящие пути спинного мозга, III: участки происхождения кортикоспинального тракта». J. Comp. Neurol
.
296
(4): 559–583. Дои:10.1002 / cne.902960405. - Тоошима, К. и Сакаи, Х (1982). «Точная кортикальная степень происхождения кортикоспинального тракта (CST) и количественный вклад в CST в различных цитоархитектонических областях. Исследование пероксидазы хрена на обезьянах». Й. Хирнфорш
.
23
: 257–269. - Чжан, С., Иде, Дж. С., Ли, К. С. (2012). «Функциональная связность в состоянии покоя медиальной верхней лобной коры». Цереб. Кора
.
22
(1): 99–111. Дои:10.1093 / cercor / bhr088. ЧВК 3236794. PMID 21572088.CS1 maint: несколько имен: список авторов (ссылка на сайт) - ^ аб
Пенфилд, В. и Уэлч, К. (1951). «Дополнительная моторная область коры головного мозга: клиническое и экспериментальное исследование».
AMA Arch. Neurol. Психиатрия
.
66
(3): 289–317. Дои:10.1001 / archneurpsyc.1951.02320090038004. PMID 14867993. - Вулси, C.N., Settlage, P.H., Meyer, D.R., Sencer, W., Hamuy, T.P. и Трэвис, А. (1952). «Паттерн локализации в премоторных областях и их связь с понятием премоторной зоны». Ассоциация исследований нервных и психических заболеваний
. Нью-Йорк, Нью-Йорк: Raven Press.
30
: 238–264.CS1 maint: несколько имен: список авторов (ссылка на сайт) - Гулд, Х.Дж. III, Кьюсик, К.Г., Понс, Т.П. и Kaas, J.H (1996). «Связь соединений мозолистого тела с картами электростимуляции моторики, дополнительной моторики и лобных полей глаз у совообразных обезьян». J. Comp. Neurol
.
247
(3): 297–325. Дои:10.1002 / cne.902470303. PMID 3722441.CS1 maint: несколько имен: список авторов (ссылка на сайт) - Муаккасса, К.Ф. И Стрик П.Л. (1979). «Входы лобных долей в моторную кору приматов: данные о четырех соматотопически организованных« премоторных »областях». Мозг Res
.
177
: 176–182. Дои:10.1016/0006-8993(79)90928-4. PMID 115545. - Миц, А. И Мудрый, С.П. (1987). «Соматотопическая организация дополнительной моторной области: картографирование внутрикортикальной микростимуляции». J. Neurosci
.
7
(4): 1010–1021. Дои:10.1523 / JNEUROSCI.07-04-01010.1987. - Gaymard, B, Pierrot = Deseilligny, C. и Rivaud, S (1990). «Нарушение последовательности управляемых памятью саккад после дополнительных поражений двигательной области». Анналы неврологии
.
28
(5): 622–626. Дои:10.1002 / ana.410280504.CS1 maint: несколько имен: список авторов (ссылка на сайт) - Герлофф, К., Корвелл, Б., Чен, Р., Халлет, М. и Коэн, Л.Г. (1997). «Стимуляция дополнительной моторной области человека мешает организации будущих элементов в сложных моторных последовательностях». Мозг
.
120
(9): 1587–1602. Дои:10.1093 / мозг / 120.9.1587.CS1 maint: несколько имен: список авторов (ссылка на сайт) - Дженкинс, И.Х., Брукс, Д.Дж., Никсон, П.Д., Фраковяк, Р.С. и Пассингем Р.Э. (1994). «Обучение двигательной последовательности: исследование с помощью позитронно-эмиссионной томографии». J. Neurosci
.
14
(6): 3775–3790. Дои:10.1523 / JNEUROSCI.14-06-03775.1994.CS1 maint: несколько имен: список авторов (ссылка на сайт) - Ли, Д. и Куэсси, S (2003). «Активность в дополнительной моторной области, связанная с обучением и выполнением во время последовательной зрительно-моторной задачи». J. Neurophysiol
.
89
(2): 1039–1056. Дои:10.1152 / ян.00638.2002. - Мусиаке, Х., Инасе, М. и Танджи, Дж. (1990). «Селективное кодирование моторной последовательности в дополнительной моторной области коры головного мозга обезьяны». Exp. Мозг Res
.
82
: 208–210. Дои:10.1007 / bf00230853.CS1 maint: несколько имен: список авторов (ссылка на сайт) - Шима К. и Танджи Дж. (1998). «Как дополнительные, так и дополнительные двигательные области имеют решающее значение для временной организации множественных движений». J. Neurophysiol
.
80
(6): 3247–3260. Дои:10.1152 / jn.1998.80.6.3247. PMID 9862919. - ^ аб
Роланд П.Е., Ларсен Б., Лассен Н.А. и Скинхой Э. (1980). «Дополнительная моторная зона и другие области коры в организации произвольных движений у человека».
J. Neurophysiol
.
43
(1): 118–136. Дои:10.1152 / ян.1980.43.1.118. PMID 7351547.CS1 maint: несколько имен: список авторов (ссылка на сайт) - ^ аб
Роланд П.Е., Скинхой Э., Лассен Н.А. и Ларсен Б. (1980). «Различные области коры человека в организации произвольных движений во внеличностном пространстве».
J. Neurophysiol
.
43
: 137–150. Дои:10.1152 / ян.1980.43.1.137. PMID 7351548.CS1 maint: несколько имен: список авторов (ссылка на сайт) - Бринкман, С (1981). «Повреждения в дополнительной моторной области мешают обезьяне выполнять задачу бимануальной координации». Neurosci. Латыш
.
27
(3): 267–270. Дои:10.1016/0304-3940(81)90441-9. PMID 7329632. - Серриен, Д.Дж., Стренс, Л.Х., Оливейеро, А., Браун, П. (2002). «Повторяющаяся транскраниальная магнитная стимуляция дополнительной моторной области (SMA) ухудшает бимануальный контроль движений у людей». Neurosci. Латыш
.
328
(2): 89–92. Дои:10.1016 / s0304-3940 (02) 00499-8. PMID 12133562.CS1 maint: несколько имен: список авторов (ссылка на сайт) - Холсбанд У., Мацузака Ю. и Танджи Дж. (1994). «Нейрональная активность в дополнительной, пре-дополнительной и премоторной коре приматов во время последовательных движений с внешними и внутренними инструкциями». Neurosci. Res
.
20
(2): 149–155. Дои:10.1016/0168-0102(94)90032-9. PMID 7808697.CS1 maint: несколько имен: список авторов (ссылка на сайт) - Пикард Н. и Стрик П. Л. (сентябрь 2003 г.). «Активация дополнительной моторной области (SMA) при выполнении движений под визуальным контролем». Цереб. Кора
.
13
(9): 977–986. Дои:10.1093 / cercor / 13.9.977. PMID 12902397. - Дик Л., Корнхубер (1978). «дополнительная» моторная кора в произвольных движениях пальцев человека ». Мозг Res
.
159
(2): 473–476. Дои:10.1016/0006-8993(78)90561-9. PMID 728816. - Каннингтон Р., Виндишбергер С., Дик Л., Мозер Е. (2003). «Подготовка и готовность к добровольному движению: высокопрофильное событие, связанное с фМРТ-исследованием реакции Bereitschafts-BOLD». NeuroImage
.
20
(1): 404–412. Дои:10.1016 / с1053-8119 (03) 00291-х. PMID 14527600. - Нгуен В.Т., Брейкспир М, Каннингтон Р. (2014). «Взаимные взаимодействия SMA и поясной коры поддерживают преддвигательную активность для произвольных действий». J Neurosci
.
34
(49): 16397–16407. Дои:10.1523 / jneurosci.2571-14.2014. ЧВК 6608485. PMID 25471577. - ^ аб
Грациано, М.С.А. (2008).
Интеллектуальная машина движения
. Оксфорд, Великобритания: Издательство Оксфордского университета. - Грациано, М.С.А. и Афлало, Т. (2007). «Отображение поведенческого репертуара на кортекс». Нейрон
.
56
(2): 239–251. Дои:10.1016 / j.neuron.2007.09.013. PMID 17964243.CS1 maint: несколько имен: список авторов (ссылка на сайт) - Грациано, М.С.А., Афлало, Т. и Кук, Д.Ф. (2005). «Движения рук, вызванные электростимуляцией в моторной коре головного мозга обезьян». J. Neurophysiol
.
94
(6): 4209–4223. Дои:10.1152 / ян.01303.2004. PMID 16120657.CS1 maint: несколько имен: список авторов (ссылка на сайт)
Первая и вторая сигнальные системы
Неоценима роль коры большого мозга в совершенствовании первой сигнальной системы и развитии второй. Эти понятия разработаны И.П.Павловым. Под сигнальной системой в целом понимают всю совокупность процессов нервной системы, осуществляющих восприятие, переработку информации и ответную реакцию организма. Она связывает организм с внешним миром.
Первая сигнальная система обусловливает восприятие посредством органов чувств чувственно-конкретных образов. Она является основой для образования условных рефлексов. Эта система существует как у животных, так и у человека.
В высшей нервной деятельности человека развилась надстройка в виде второй сигнальной системы. Она свойственна только человеку и проявляется словесным общением, речью, понятиями. С появлением этой сигнальной системы стали возможными отвлеченное мышление, обобщение бесчисленных сигналов первой сигнальной системы. По И.П.Павлову, слова превратились в «сигналы сигналов».
Возникновение второй сигнальной системы стало возможным благодаря сложным трудовым взаимоотношениям между людьми, так как эта система является средством общения, коллективного труда. Словесное общение не развивается вне общества. Вторая сигнальная система породила отвлеченное (абстрактное) мышление, письмо, чтение, счет.
Слова воспринимаются и животными, но совершенно отлично от людей. Они воспринимают их как звуки, а не их смысловое значение, как люди. Следовательно, у животных нет второй сигнальной системы. Обе сигнальные системы человека взаимосвязаны. Они организуют поведение человека в широком смысле слова. Причем вторая изменила первую сигнальную систему, так как реакции первой стали в значительной мере зависеть от социальной среды. Человек стал в состоянии управлять своими безусловными рефлексами, инстинктами, т.е. первой сигнальной системой.
Функции коры мозга
Знакомство с наиболее важными физиологическими функциями коры большого мозга свидетельствует о необычайном ее значении в жизнедеятельности. Кора вместе с ближайшими к ней подкорковыми образованиями является отделом центральной нервной системы животных и человека.
Функции коры головного мозга — осуществление сложных рефлекторных реакций, составляющих основу высшей нервной деятельности (поведения) человека. Не случайно у него она получила наибольшее развитие. Исключительным свойством коры являются сознание (мышление, память), вторая сигнальная система (речь), высокая организация труда и жизни в целом.
дальнейшее чтение
- Принципы неврологии (2000), 4-е изд., Kandel et al.
- Дебаэр, Ф., Вендерот, Н., Сунаерт, С., Ван-Хек, П., Суиннен, С. П. (июль 2003 г.). «Внутренняя и внешняя генерация движений: дифференциальные нервные пути, участвующие в бимануальной координации, выполняемые при наличии или отсутствии усиленной визуальной обратной связи». NeuroImage
.
19
(3): 764–76. Дои:10.1016 / с1053-8119 (03) 00148-4. PMID 12880805.CS1 maint: несколько имен: список авторов (ссылка на сайт) - Воробьев; и другие. (1998). «Разделение мезиальной области человека 6: цитоархитектонические свидетельства для трех отдельных областей». Eur J Neurosci
.
10
(6): 2199–203. Дои:10.1046 / j.1460-9568.1998.00236.x. PMID 9753106.